MAJOR GROUPS CONSIDERED
Amebae
Amebae are protozoa generally characterized as using pseudopodia as organelles of motility (Sleigh 1991). Most amebae now are included in the Supergroup Amoebozoa; a few other species having an ameba-like stage (e.g., Dientamoeba, Naegleria) are classified in the Supergroup Excavata (Adl et al.
2005) (App. 1: Table 5). A trophozoite is any protozoan in an asexual phase of development; among amebae the trophozoite is the active feeding stage. For intestinal amebae, the trophozoite will form a cyst stage that is well suited to withstand environmental conditions outside of the host, prior to being passed out with feces.Amoebozoa are characterized by ameboid locomotion generally, with non-eruptive, morphologically variable pseudopodia (lobopodia). The cells are “naked” or testate; cells usually are uninucleate, rarely binucleate or multinucleate. Cysts are common, but can be morphologically variable among species (Adl et al. 2005). Within the amebae, there are several groups that can cause diseases among vertebrates; of these, the genus Entamoeba is the most important. Entamoeba spp. lack mitochondria, have a simple endomembrane system, and undergo mitosis that is closed with an internal spindle (Sleigh 1991, Adl et al. 2005). Trophozoites have one nucleus. Cyst stages typically have four nuclei (Fig. 6.1), although immature cysts may have only one of two nuclei (Leber and Novak 1999). Most parasitic amebae inhabit the large intestine; cyst formation is a chemicaldependent process in the colon (Eichinger 2001).
Entamoeba histolytica also has been reported from nonhuman primates and occasionally canines (Kocan 2001a). This parasite can be found worldwide but is more prevalent in tropical and subtropical regions (Leber and Novak 1999). Humans are considered the primary reservoir for Entamoeba histolytica (Leber and Novak 1999), and this parasite is the third most common cause of parasite-induced deaths among humans (Martinez-Paloma 1993).
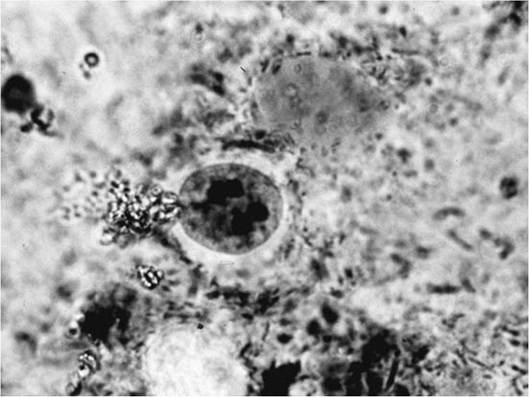
FIGURE 6.1 Entamoeba spp.
multinuclear cyst.
Entamoeba invadens is an important parasite of reptiles (Frank 1976, Lucius and Loos-Frank 1997); it infects carnivorous reptiles such as snakes, where it causes disease, and herbivorous reptiles such as turtles, in which it lives as a commensal (Ratcliffe and Geiman 1938, Meerovitch 1958, Jakob and Wesemeier 1995). Entamoeba spp. also have been reported in waterfowl (Quortrup and Shillinger 1941).
Opportunistic, free-living amebae that can act as opportunistic parasites include Balamuthia spp. and Acanthamoeba spp.; members of both genera have a cyst stage. Acanthamoeba spp. and Balamuthia spp. inhabit the neurological tissues of infected hosts (Martinez and Visvesvara 1997, John 2001). No animal reservoir for these potentially pathogenic free-living amebae has been identified (John 2001). Interestingly, Acanthamoeba castellanii may play a role in the maintenance of pathogenic bacteria and fungi in the environment (Essig et al. 1997, Abd et al. 2003, Steenbergen et al. 2004).
The life cycle of the ameba within the Amoebozoa is simple and direct. Division among ameba trophozoites is by simple fission. In cyst formation, trophozoites typically empty their food vacuoles, round up, get a little smaller, and surround themselves with a thin, delicate wall that is relatively sensitive to desiccation but fairly resistant to many chemicals. Some nuclear multiplication typically occurs in the phase, and on infecting a new host, the cyst opens (excystment) in the small intestine before the resulting amebae migrate down to the large intestine. Some cytoplasm is associated with each nucleus in the cyst after excystment.
Apicomplexa (Sporozoa)
Apicomplexa are single-celled protists named for an apical complex of organelles in the infective stages (sporozoites, merozoites) that is unique among living organisms (Atkinson and van Riper 1991, Sleigh 1991).
Historically called Sporozoa (“spore animals”), these protists currently are classified in the Supergroup Chromalveolata (App. 1: Table 5) (Adl et al. 2005). Apicomplexa produce spores at some time during their life cycle. While Apicomplexa generally are characterized by the absence of obvious external motility organelles (Bush et al. 2001), locomotion can occur by gliding, body flexion, longitudinal ridges, and/or flagella in some stages (Adl et al. 2005). Virtually all forms are parasitic (Adl et al. 2005).The life cycles of Apicomplexa are complex and include both direct (monoxenous) and
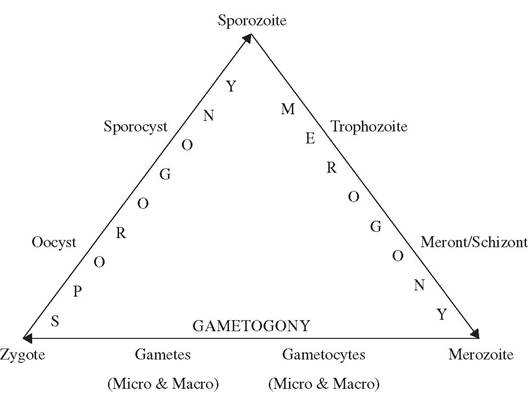
FIGURE 6.2 Generalized life
cycle for the Apicomplexa.
indirect (heteroxenous) life cycles; all include both sexual and asexual phases (Bush et al. 2001). We use the terms and descriptions of Baer, as modified by Olsen and Bush (Baer 1951, 1971; Olsen 1974; Bush et al. 2001) to present the basic cycle. Although complex, the same terms and processes apply to virtually all Apicomplexa life cycles.
Three distinct processes of multiplication generally occur among the Apicomplexa, including one sexual and two asexual forms (Fig. 6.2). Sexual reproduction (gametogony) among Apicomplexa, where known, is by syngamy (“gamete fusion”) followed by immediate meiosis to produce haploid progeny. Some authors prefer the term “gamogony” rather than “gametogony” (Duszynski and Upton 2001). Asexual reproduction of haploid stages occurs by binary fission, merogony (a form of multiple fission also called schizogony in earlier literature), endodyogeny (binary fission within a cell), and/or endopolyogeny (multiple fission within a cell) (Adl et al. 2005). If the final products of multiplication are merozoites, the process is called merogony, and if sporozoites, it is called sporogony (Bush et al. 2001). Merogony, often called schizogony in earlier literature, is a form of multiple fission leading to the formation of merozoites.
Sporogony is a form of binary fission in the spore stage, leading to development of sporozoites.Sporozoites are the infective stage for vertebrate hosts and, once in the proper vertebrate host, infect the host cells and develop into trophozoites. Through repeated division (merog- ony), trophozoites form meronts (earlier called schizonts), which further develop into mero- zoites. For some Apicomplexa, merozoites may function as trophozoites by reinvading the same kind of host cell and producing additional generations of merozoites (Bush et al. 2001). Eventually, merozoites invade new vertebrate host cells and differentiate into either micro - or macrogametes through gametogony; in many cases the merozoite may first undergo repeated division to form many microgametes. Eventually, the micro - and macrogametes fuse to form a diploid zygote, followed by repeated meiotic division of this zygote to form oocysts, a protective stage within which one or more sporocysts develop, with each sporocyst further forming one of more sporozoites within itself. There are numerous variations as to the hosts and host tissues within which these various stages occur, whether the parasite has a direct life cycle or requires intermediate hosts or vectors, but the basic life cycle stages and three multiplication processes retain considerable consistency among the Apicomplexa. Although the taxonomy of the Apicomplexa frequently has been revised, three common groups of concern for wildlife diseases include
the coccidia, malaria parasites, and piroplasms (App. ι: Table 5) (Cox 1991, Patterson 2000, Adl et al. 2005).
Coccidia
Coccidia, variously classified as Eimeriida (Cox 1991), Eimeriorina (Bush et al. 2001), or Coccidiasina (Adl et al. 2005) are relatively widespread among birds and mammals. Every species of vertebrate ever examined intensively over a broad geographic range has at least one coccidian species unique to it, and some may have many more species (Duszynski and Upton 2001).
Thus, there are many different species of coccidia causing specific problems, but very few species of broad impact in wildlife diseases.There are at least 15 genera of coccidia from birds and mammals (Todd and Hammond 1971, Lindsay and Todd 1993, Long 1993, Cole 1999a, Friend and Franson 1999). The focus in this text primarily will be on the genera Eimeria, Toxoplasma, and Sarcocystis. However, others, including Hepatozoon spp. (Craig 2001) and Besnoitia spp. (Leighton and Gajadhar 2001), can be of importance as well.
Oocyst morphology is an important means by which various members of the coccidia can be identified, particularly with the numbers and combinations of sporocysts and sporozoites within each oocyst (Bush et al. 2001). For example, oocysts of members of Eimeria have four sporocysts, each of which has two sporozoites; in contrast, members of the genus Isos- pora have two sporocysts, each of which has four sporozoites.
Among coccidia (e.g., Eimeria, Cryptosporidium, Cyclospora) with a monoxenous life cycle (Fig. 6.3), the infective sporozoite typically is ingested from contaminated soil or water and inhabits the intestine or liver, leaving again as undeveloped oocysts. Once out of the host, the oocysts sporulate to develop sporocysts and sporozoites in the ambient environment, and the sporozoites become infectious for a new host. Monoxenous coccidia most commonly are reported among herbivorous hosts.
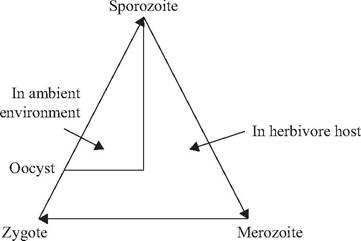
FIGURE 6.3 Generalized direct (monoxenous) life cycle for coccidia.
Among coccidia (e.g., Sarcocystis, Toxoplasma) with a heteroxenous life cycle (Fig. 6.4), the sporozoite is ingested by the intermediate vertebrate host and develops to a trophozoite stage. When the intermediate host is eaten by the predatory definitive host, the trophozoite stage develops further to the oocyst stage, which is shed into the ambient environment. Following sporulation, the life cycle is completed when oocysts containing infective sporozoites are ingested by a susceptible host.
Heteroxenous coccidia typically use carnivores as definitive hosts and herbivorous prey species as intermediate hosts. A few coccidian parasites (e.g., Isospora) can use optional (paratenic) vertebrate hosts (Dubey 1993). In paratenic hosts, the ingested sporozoite may become encysted (unizoite cyst) before being ingested by the predatory definitive host (Dubey 1993).There is evidence for an evolutionary transition among coccidia genera from monox- enous to heteroxenous life cycles (Belova and Krylov 2003, Slapeta et al. 2003). Genera such as Sarcocystis, Frenkelia, and Besnoitia parasitize carnivores as definitive hosts and have required intermediate hosts (Dubey
1993). The closely related genus Toxoplasma functions as a transitional form; while it can complete a one-host life cycle in the cat, it primarily uses intermediate hosts to facilitate completion of its life cycle (Sanger 1971, Jindrichova et al. 1975, Dubey and Odening 2001). Some members of the genus Isospora
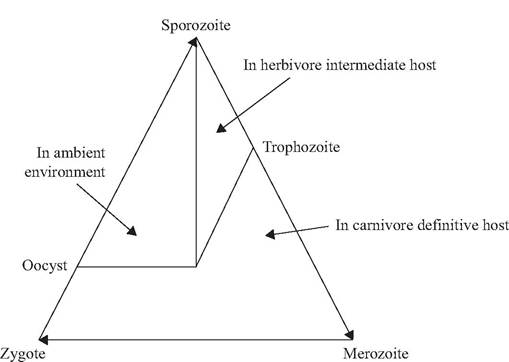
FIGURE 6.4 Generalized indirect (heteroxenous) life cycle for coccidia.
also have this pattern (Dubey 1993). Details of these evolutionary relations are incomplete (Slapeta et al. 2003).
Malaria Parasites (Haemosporidia)
Malaria parasites are blood parasites found in a wide variety of mammals, birds, and reptiles; all are transmitted by hematophagous arthropods. The term “malaria” stems from earlier beliefs that bad air or gases (“mal-air”) from swamps caused the disease in humans. Variously identified as Haemosporina (Atkinson and van Riper 1991), Haemospororina (Bush et al. 2001), and Haemospororida (Cox 1991, Adl et al. 2005), all malarial parasites are vector- borne and involve a hematophagous invertebrate. The prefix “haemo” is the Greek root for blood, and “sporo” is the root for spore or seed.
Genera of importance to wildlife include Haemoproteus, Leucocytozoon, and Plasmodium among birds (Atkinson and van Riper 1991, van Riper et al. 1994) and, to a lesser extent, Plasmodium in reptiles, rodents, and primates (Collins and Aikawa 1993, Cox 1993). Two genera causing avian malaria each have a large number of species, with each species often parasitizing a relatively limited host range; Leucocytozoon has 60 and Haemoproteus has 133 species (Bennett et al. 1994). In contrast, Plasmodium spp. generally have broader host specificity (Bennett et al. 1982). There are about 38 species of Plasmodium described from mammals (Collins and Aikawa 1993, Cox 1993, Lopez-Antunano and Schmunis 1993), 34 species from birds (Bennett et al. 1993), and 87 species from reptiles (Telford 1994).
Malarial parasites have been recorded in about 68% of the avian species examined (Atkinson and van Riper 1991). Haemoproteus is the most common genus and has been reported in about 67% of malaria-infected birds, followed by Plasmodium (42%) and Leucocytozoon (39%) (Atkinson and van Riper 1991). Some host families (e.g., Fringillidae, Columbidae, Phasianidae) have relatively high prevalences of infection, whereas others (e.g., Laridae, Scolopacidae, Charadriidae) are only rarely infected with blood parasites (Atkinson and van Riper 1991). The reasons for these differences may involve behavior, habitat, suitable vectors, climate, environmental conditions, and host specificities (Atkinson and van Riper 1991).
Malarial parasite life cycles involve asexual reproduction in both the vertebrate host and arthropod vector, and sexual reproduction in the arthropod (Fig. 6.5) (van Riper et al. 1994). The arthropod hosts for Haemoproteus are members of the Family Hippoboscidae for upland birds and members of Culicoides
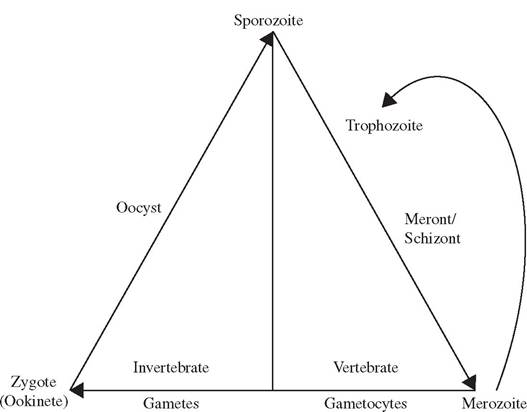
FIGURE 6.5 Generalized malarial life cycle.
for waterfowl (Bush et al. 2001). Members of Leucocytozoon are transmitted by black flies (Family Simuliidae) (Cook 1971b). Among Plasmodium spp., mosquitoes of the genus Anopheles transmit all species infecting mammals as well as a few bird malarias; however, species of Culex transmit most avian malarias of Plasmodium spp. (van Riper et al. 1994).
The invertebrate is the definitive host because it is the site of sexual reproduction and thus is where the adult stage of the parasite occurs. In contrast, the vertebrate is the intermediate host since only asexual reproduction occurs within it. The infective sporozoite stage enters the susceptible vertebrate host with salivary gland secretions during the bite of the hematophagous vector (Atkinson and van Riper 1991). After invading the host and undergoing intracellular growth, development, and asexual reproduction (merogony) in the host tissues, merozoites are formed. Merozoites invade circulating blood cells and may undergo additional merogony (e.g., Plasmodium) or undergo gametogony to form male (micro-) or female (macro-) gametocytes that are infective for another susceptible hematophagous vector (Atkinson and van Riper 1991).
Once ingested by the invertebrate host, gametocytes form gametes that fuse in the arthropod midgut to form motile zygotes (ookinetes), which, in turn, penetrate the midgut epithelial cells (Atkinson and van Riper 1991). There, ookinetes round up into oocysts and undergo merogony to produce numerous sporozoites. Sporozoites rupture from the oocysts into the hemocoel of the vector and move to the salivary glands for injection into the next susceptible host (Atkinson and van Riper 1991).
While similar, the life cycles of Plasmodium, Leucocytozoon, and Haemoproteus differ in the vertebrate host tissues they use for schizogony. Leucocytozoon and Haemoproteus undergo schizogony only in fixed, non-circulating cells in the host such as hepatocytes (liver cells) and vascular endothelial cells (cells lining blood vessels); further, only the gametocyte stage is observed in the circulating blood for these genera (Atkinson and van Riper 19 91). In contrast, Plasmodium undergoes merogony in both fixed non-circulating cells (exoeryth- rocytic merogony) and circulating blood cells (erythrocytic merogony); thus, both gametocytes and meronts may occur in erythrocytes (Atkinson and van Riper 1991). The occurrence
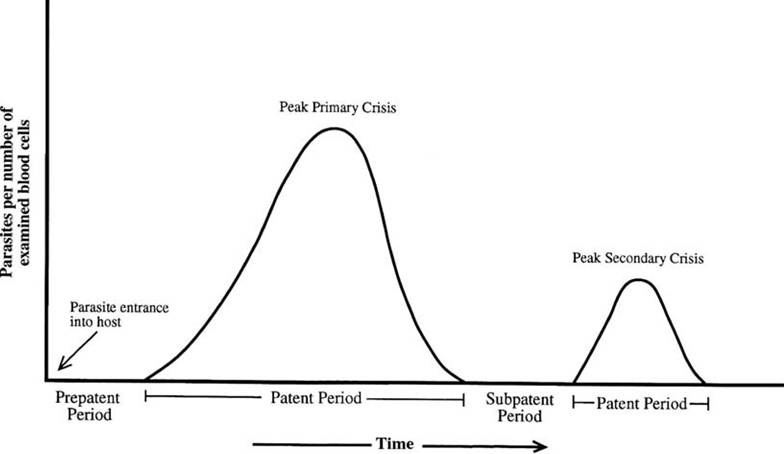
FIGURE 6.6 Description of a malarial infection (from Parasitic protozoa, Volume 7, C. van Riper III et al., Plasmodia of birds, pp. 73-140, J. Kreier (ed.), Copyright Academic Press 1994).
of erythrocytic meronts is a key difference of Plasmodium from Leucocytozoon and Haemoproteus (Atkinson and van Riper 1991).
A distinct vocabulary is associated with malarial infections. The occurrence of parasites in the blood is termed “parasitemia” (Fig. 6.6). Following the bite of an infected hematophagous vector, the period of time between initial infection and when parasites are detected in the blood is termed the prepatent period; this is the time during which parasites are developing in various host tissues before invading the blood (Atkinson and van Riper 1991). The patent period is defined as the time when parasites can be observed in the blood, generally with a thick blood smear. The acute phase is when parasites first appear in the blood and rapidly increase in number. The crisis phase occurs when the parasitemia in circulating blood and the resulting physiological stresses reach a peak. Following the patent period, a latent or subpatent phase occurs when immune responses have reduced the parasitemia to low levels, where the parasites no longer are detectable in blood smears, and the surviving hosts have few or no overt signs of infection (Atkinson and van Riper 1991, van Riper et al. 1994). For species typical of temperate areas, a relapse of parasites in the vertebrate host's blood typically occurs the following spring, and serves to infect the new generation of hematophagous vectors.
While some species of Haemoproteus sometimes are associated with host pathology (Julian and Galt 1980, Atkinson et al. 1988, Cannell et al. 2013), many species have little or no pathogenic effect (Cook 1971a, Bennett et al. 1988). Plasmodium and Leucocytozoon more commonly are associated with diseases among infected hosts (Atkinson 1999).
Piroplasms
Piroplasms are sporozoan parasites of mammals and birds passed by ticks of the Family Ixodidae (Allan 2001, Kocan and Waldrup 2001). All cause a hemolytic anemia in the vertebrate host. Two families are recognized: Babesidae and Theileridae.
At least 73 species of Babesia have been reported from mammals (Kakoma and Mehlhorn 1994), as well as 13 species from birds (Peirce 2000), and several from reptiles (Bush et al. 2001). Among mammals, species of
Babesia infect primarily cervids and bovids, as well as some carnivores and rodents around the world. The 13 recognized species of Theileria all occur in mammals, primarily wild and domestic ungulates (Mehlhorn et al. 1994). The taxonomic status of the genus Cytauxzoon currently is unclear, as some scholars combine it with Theileria (Mehlhorn et al. 1994) and others keep it separate (Kocan and Waldrup 2001). Species of Theileria and Cytauxzoon occur in a variety of mammals throughout the world (Barnett and Brocklesby 1969), Theileria cervi, T. annulata, and Cytauxzoon felis are known from wild mammals of North America (Kocan and Waldrup 2001).
The vectors for Babesia spp. have been identified for only 17 of the 73 recognized species; all are ticks (Kakoma and Mehlhorn 1994). Known ixodid tick vectors for Babesia include species of Boophilus, Dermacentor, Haemaphy- salis, Rhipicephalus, and Hyalomma (Kakoma and Mehlhorn 1994). For Theileria, known ixo- did tick vectors include species of Rhipicepha- lus, Hyalomma, and Amblyomma (Mehlhorn et al. 1994).
The life cycles of piroplasms involve processes similar to those of malaria parasites (Fig. 6.5). Merogony occurs in the vertebrate host, whereas gamete maturation, fusion, and formation of a motile zygote (kinete) occur in the tick intestine (Bush et al. 2001).
Piroplasm sporozoites are injected into susceptible hosts with the saliva of infected ixodid ticks (Kakoma and Mehlhorn 1994, Mehlhorn et al. 1994). Babesia spp. sporozoites invade the cytoplasm of erythrocytes of the vertebrate hosts and undergo several cycles of merogony to form merozoites (Kakoma and Mehlhorn
1994). Theileria spp. sporozoites invade the cytoplasm of erythrocytes, as well as newly dividing lymphocytes, which continue to divide together with the parasite (Mehlhorn et al. 1994, Bush et al. 2001).
Sexual reproduction occurs in the tick vector (Mackenstedt et al. 1990). For both genera, the newly formed merozoites invade other erythrocytes and are then ingested by the appropriate tick vectors, in which they undergo gamogony (gametogony) and form gametes and kinetes (motile zygotes) in the tick gut. Babesia spp. kinetes can penetrate a number of tick cell types and divide by multiple fission to produce additional kinetes (Kakoma and Mehlhorn 1994). Kinetes eventually migrate to the tick salivary glands, where sporogony results in the production of many sporozoites, which, in turn, are transmitted to the next generation of vertebrate hosts (Bush et al. 2001). For Theile- ria, kinetes develop only in the salivary glands of the tick hosts (Mehlhorn et al. 1994).
Members of the Theileridae and Babesi- dae differ in a number of ways. Theileridae undergo merogony in lymphocytic cells in addition to their merogony in erythrocytes; in contrast, members of the Babesidae generally undergo asexual multiplication (merogony) only in erythrocytes of the vertebrate host. However at least two species of Babesia also may invade lymphocytes as sporozoites (Moltmann et al. 1983, Mehlhorn et al. 1986, Kakoma and Mehlhorn 1994). Members of the Babesidae undergo both transstadial (across stages of the tick vector life cycle) and transovarial transmission in the tick vector; in contrast, only trans- stadial transmission is known to occur in the Theileridae (Kocan and Waldrup 2001). Thus, transmission of Theileria requires multi-host ticks whereas Babesia, which can be transmitted transovarially, can be transmitted by ł-host ticks (Mehlhorn et al. 1994). For Babesia, ticks are biological vectors; mechanical transmission by inoculation of infective blood by flying insects does not occur (Kakoma and Mehlhorn 1994). In contrast to Babesia, Theileria also can be transmitted mechanically by the bite of bloodsucking insects (Mehlhorn et al. 1994).
Ciliophora (Ciliates)
The Ciliophora (ciliates) are a large and diverse group of protista related to the Api- complexa, which are also in the Alveolata (App. 1: Table 5) (Adl et al. 2005). All members have cilia as organelles of motility; species are distinguished on the basis of their pattern of cilia and associated cortical structure, and on the basis of their nuclear structure and function (Bush et al. 2001). Ciliates commonly divide by binary fission, although conjugation can occur (Bush et al. 2001). In contrast to the Apicomplexa, there are very few ciliates that cause disease among vertebrates (Bush et al. 2001). All ciliates have a direct (monox- enous) life cycle. Among wildlife, only Balantidium coli has been reported to cause disease with any regularity (Lucius and Loos-Frank 1997, Kocan 2001a). Some ciliates, including B. coli, produce resistant cyst stages (Bush et al. 2001).
Excavata (Flagellates)
A diverse variety of eukaryotic single-celled organisms have a life history stage using one or more flagellae as organelles of motility. Parasitic f lagellates infect most animal phyla and occupy a variety of host habitats, including the intestines, reproductive tract, deep body tissues, as well as intracellularly and extracellularly in the blood vascular system (Bush et al. 2001).
Until recently, flagellates were believed to be closely related to amebae (Bush et al. 2001). However, most flagellates currently are classified within the Supergroup Excavata (Adl et al. 2005). Further, within the Excavata, flagellates are broadly distributed among three of the six First Rank subgroups, including Fornicata, Parabasilia, and Euglenozoa. From a clinical perspective, flagellates often have been distinguished as intestinal flagellates (included in Fornicata and Parabasilia) or hemoflagellates (in Euglenozoa) (Chandler and Read 1961, Kocan 2001a).
Intestinal Flagellates
Most intestinal flagellates of concern to wildlife fall within the Trichomonadida subgroup of the Parabasilia (Adl et al. 2005). Important genera include Histomonas and Trichomonas. Their flagellae are often associated with a lamellar undulating membrane. Another member of this group, Dientamoeba fragilis, no longer has a flagellum (BonDurant and Wakenell 1994).
Most trichomonad species are not established pathogens for mammals or birds (BonDurant and Honigberg 1994). However, a few are of importance, including Histomonas meleagridis and Trichomonas gallinae in birds (BonDurant and Honigberg 1994, BonDurant and Wakenell 1994).
Another group among the Excavata, the For- nicata, lack typical mitochondria and generally have either a single nucleus or pair of nuclei (Adl et al. 2005). Giardia is one important genus in this group, infecting amphibians, reptiles, birds, and mammals, including humans (Olson and Buret 2001). Giardia spp. have two nuclei in their trophozoites and four nuclei in their cysts (Garcia and Bruckner 1997). They lack a functional feeding apparatus; each organism has one posteriorly directed flagellum running through the length of the cell axially and within the cytoplasm (Adl et al. 2005). While classification of Giardia species still is quite controversial (Kulda and Nohynkova 1994), members of the genus cause serious diarrheal disease in humans as well as diarrhea and allergies in domestic animals; Giardia also infects a variety of wild animals, but usually with fewer clinical signs (Olson and Buret 2001).
Life cycles among the intestinal flagellates are varied, but most generally are simple. The intestinal flagellates divide by binary fission, and most, including Giardia and Trichomonas, have a direct life cycle; Giardia forms cysts, whereas Trichomonas and Histomonas lack cyst stages (BonDurant and Honigberg 1994, BonDurant and Wakenell 1994, Kulda and Nohynkova 1994).
Transmission is direct, including ingestion of trophozoites (e.g., Trichomonas gallinae) or encysted stages (e.g., Giardia), or sexual intercourse (e.g., Trichomonas vaginalis). Rarely, transmission also may be indirect, carried through helminth infections; both Histomonas meleagri- dis (Graybill and Smith 1920) and Dientamoeba fragilis (Burrows and Swerdlow 1956, Ockert and Schmidt 1976, Sukanahaketu 1977) are
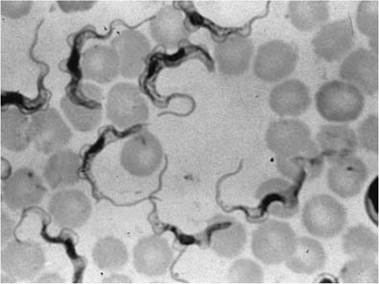
FIGURE 6.7 Trypanosoma evansi in mouse blood (Courtesy of W. Frank, Universitat Hohenheim, Germany).
helminth-transmitted parasites. For H. meleagri- dis, the nematode Heterakis gallinae is critical to the natural transmission of the parasite; earthworms further serve as paratenic hosts in this complex (BonDurant and Wakenell 1994). For D. fragilis, the human pinworm Enterobius ver- micularis plays an important role in the life cycle.
Hemoflagellates
Among the Excavata, the Euglenozoa include parasitic hemoflagellates of importance to wildlife (Adl et al. 2005). Two important parasite genera in this group, Trypanosoma and Leishma- nia, both have a single flagellum; they multiply by binary fission and only rarely undergo sexual reproduction (Bush et al. 2001, Adl et al. 2005). These hemoflagellates live in the blood, lymph, and other tissues of vertebrates (Fig. 6.7). Morphological features such as length, shape, and location within the parasite of organelles such as the basal body, kinetoplast, nucleus, and flagellum are used to distinguish various species (Logan-Henfrey et al. 1992).
Most hemoflagellates are found in tropical and subtropical regions of the world. Species of Leishmania are found in Central and South America, Asia, India, and China, with occasional reports in North America (Lainson 1982). These parasites live in the macrophages of the lymph nodes, spleen, and liver of their vertebrate hosts and are parasites of mammals; carnivores and rodents commonly serve as important vertebrate reservoirs. The invertebrate vectors are sand flies (Phlebotomus spp.) (Lainson 1982).
Species of Trypanosoma infect a wide variety of vertebrates and arthropods in Africa, Asia, and North and South America (Wells and Lumsden 1969). Although most wildlife-related studies with Trypanosoma have involved mammals (Molyneux 1982), all vertebrate classes are parasitized by species of Trypanosoma (Herman 1969, Bardsley and Harmsen 1973, Baker 1976, Lom 1979, Telford
1995). Avian hosts include raptors, waterfowl, columbiforms, galliforms, and passerine birds (Gylstorff* and Grimm 1987).
The taxonomy of Trypanosoma spp. is controversial. Members of the genus historically have been divided into a) species that develop in the midgut or hindgut of their insect vectors and are passed by scratching infective stages from contaminated feces into a wound produced by the bite of an insect vector (Stercoraria), and b) species that develop in the midgut or mouth parts of their insect vectors and are passed directly by arthropod bite (Salivaria) (Hoare 1966). In more recent years, parasites in the genus Trypanosoma have been viewed as a complex taxonomic unit that likely includes many different genera; species designations often include reference to a new proposed genus, as in Trypanosoma (Schizotrypanum) cruzi, T. (Trypanozoon) brucei, T. (Nannomonas) congolense, and T. (Duttonella) vivax (Bush et al. 2001). However, we continue using the standard Trypanosoma genus in this text.
Trypanosoma cruzi, a Stercoraria, is a significant human problem in Central and South America; it develops in the hind-gut of its hosts and infects its vertebrate host through the vector's excrement, which is rubbed into the bite wound by the host (Acha and Szyfres 2003). About 150 mammal species can serve as vertebrate hosts, ranging from marsupials (Didel- phis marsupialis) to primates; prevalence can be high in cats, dogs, rodents, and both domestic and wild lagomorphs (Acha and Szyfres 2003). About 100 species of triatomid bugs (Hemip- tera: Reduviidae) can transmit these parasites,
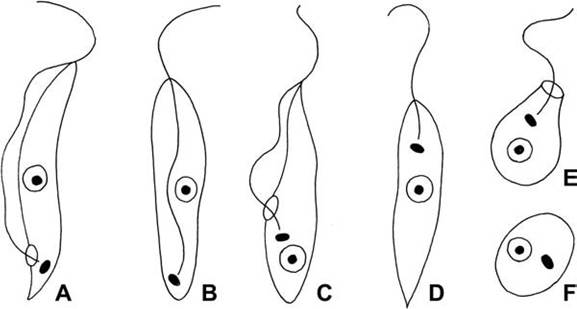
FIGURE 6.8 Key developmental stages of hemoflagellates. a. trypomastigote; b. Opisthomastigote; c. epimastigote; d. promastigote; e. choanomastigote, f amastigote (Lucius and Loos-Frank, 1997; courtesy R. Lucius, Humboldt University Berlin).
but only about three are considered as significant vectors for human disease (Sousa and Johnson 1973, Cedillos 1975, Acha and Szyfres 2003).
Trypanosomiasis in Africa is one of the most important diseases of humans and livestock (Wells and Lumsden 1969, Logan- Henfrey et al. 1992). Wildlife serve as reservoir hosts (Kocan 2001b). Trypanosoma brucei is one important species; two subspecies, T. brucei gambiense and T. brucei rhodesiense, are important causes of sleeping sickness among humans (Wells and Lumsden 1969). Another subspecies, T. brucei brucei, along with T. vivax and T. congolense, are important parasites, causing considerable morbidity and mortality with a disease called nagana among cattle of Africa (Wells and Lumsden 1969). All of these use species of tsetse flies (Glossina spp.) as vectors (Wells and Lumsden 1969).
Trypanosoma and Leishmania have relatively complex life cycles involving both vertebrates and hematophagous insects (Lucius and Loos- Frank 1997, Bush et al. 2001). Several distinct life history stages occur among the vertebrate and invertebrate hosts (Hoare and Wallace 1966) (Fig. 6.8). The names of the life cycle stages each are derived from a combination of the Greek root “mastigote,” meaning whip, and an appropriate prefix (Hoare and Wallace 1966). The main morphological features distinguishing the developmental stages in trypanosomes are the position of the kinetoplast (a specialized region of the mitochondrion containing a dense network of DNA), the way in which the flagellum is attached to the body of the cell, and the extent of the flagellum’s extension anteriorly (Logan-Henfrey et al. 1992).
When present, the trypomastigote and amastigote forms typically develop in vertebrates, and the epimastigote and promastigote develop in the intestinal tract of the infected insects (Lucius and Loos-Frank 1997). The trypomastigote stage is the common form in the peripheral blood of the vertebrate host during early stages of infection for Trypanosoma (Bush et al. 2001, Kocan 2001b). “Metacyclic trypomastigote” is a term for the stage in the invertebrate host that is infective for the susceptible vertebrate host. Amastigotes are stages of some trypanosomes infecting vertebrate tissues (Bush et al. 2001). Promastigotes are stages typically found in the midgut of infected insects for Leishmania and some Trypanosoma. In Trypanosoma, the epimastigote stages occur in the insect midgut; epimastigotes divide by binary fission (Bush et al. 2001). The sphaero- mastigote (not pictured) is a stage identified in the T. cruzi life cycle as occurring between amastigote and epimastigotes stages in mammalian hosts (Bush et al. 2001).
Collectively, metacyclic stages are defined as the final stage of development of parasitic
TABLE 6.1
Life History Types Found among Protista, with Representative Genera for Each
| Life History Type | Representative Genera |
| Direct (Homoxenous) Life Cycles | |
| No cyst stage | Trichomonas |
| Cyst stage | Entamoeba, Giardia, Eimeria, Balantidium |
| May use paratenic host | Isospora |
| Indirect (Heteroxenous) Life Cycles | |
| Vertebrate Intermediate host | Toxoplasma, Sarcocystis |
| Arthropod-borne | Plasmodium, Leucocytozoon, Babesia, Hepatocystis, Trypanosoma, Leishmania |
| Helminth-borne | Histomonas, Dientamoeba |
| Free-living Soil and Water Protista | |
| (Opportunistic parasites) | Naegleria, Acanthamoeba, Balamuthia |
kinetoplastid flagellates during the invertebrate (or vector) phase of their life cycle and are the infective stages for their vertebrate host (Bush et al. 2001). In T. cruzi, it is the infective stage that develops in the reduviid rectum from epimastigotes. In Salivaria, the infective (trypomastigote) stage develops in the mouthparts of the insect (Bush et al. 2001); these stages do not divide.
The trypomastigote and epimastigote stages are found only in the genus Trypanosoma; in contrast, the amastigote and promastigote stages are represented in some species among both Trypanosoma and Leishmania (Bush et al. 2001). Trypomastigote stages occur among all members of Trypanosoma; amastigotes are tissue stages found among the infected vertebrates with T. cruzi, Leishmania spp., and others. A procyclic stage refers to any developmental stage in the life cycle of a hemoflagellate in the invertebrate host. These most often are found in the midgut of the host (Bush et al. 2001).
The role of wildlife is complex for these parasites. Most infections by hemoflagellates in wildlife are benign (Baker 1969, Kocan 2001b), but some infecting humans and domestic animals can cause severe diseases (Molyneux 1982, Bruckner and Labarca 1999). In southern
Mexico, and in Central and South America, Trypanosoma cruzi is a significant cause of morbidity and mortality among humans (Chagas' disease) (Acha and Szyfres 2003). There are not many records of trypanosomes being pathogenic to African ungulates (Baker 1969); this could be due to tolerance of these vertebrates to the parasites or to predation of any unfit animal (Bertram 1973, Carmichael and Hobday 1975, Molyneux 1982). Based on laboratory experiments with captive African ungulates, some animals have lesions but generally appear to be unaffected (Molyneux 1982).
More on the topic MAJOR GROUPS CONSIDERED:
- APPENDIX ONE Systematics of Major Parasites Groups
- Lameness is not only a major economic problem, but it is also a major welfare issue - for both the cow and the herdsman!
- After years of promoting savings groups, I have identified nine principles that explain why the number of savings groups has grown so quickly,
- Prima Facie vs. all Things Considered
- Who was Ivan Mazepa, and why is he considered a “traitor” in Russia?
- ADDITIONAL ELEMENTS CONSIDERED IN CREDIT ENHANCEMENTS
- Two Types of Groups
- India's Self-Help Groups
- Savings Groups as Platforms for Innovation
- Narrative Construction of Individuals and Groups
- Savings Groups: Turning Banking on Its Head
- The Major Festivals
- Since the discovery of the principles of inheritance (Gregor Mendel, 1865), genetic composition of a cell is considered as the blueprint of life.