LIFE HISTORY TYPES AMONG PROTOZOA
There is a wide variety of life history strategies used by parasitic protozoa to be successful. The major ones of importance to wildlife are presented in this section (Table 6.1).
Direct (Homoxenous) Life Cycles
No Cyst Stage
TRICHOMONAS CALLINAE
CAUSATIVE AGENT (CLASSIFICATION, morphology) The genus Trichomonas and other trichomonads are classified among the Parabasilia of the Supergroup Excavata (App. 1: Table 5) (Adl et al. 2005). The genus Trichomonas includes a number of species infecting birds and mammals, of which one, Trichomonas gallinae, is a wildlife pathogen that infects the upper respiratory tract of birds (Cole 1999b). Another, T. foetus, is a parasite of the urogenital system of cattle (BonDurant and Honigberg 1994), and T. vaginalis infects the urogenital system of humans (Leber and Novak 1999).
host range and distribution Trichomonas gallinae has a worldwide distribution, with the domestic pigeon (Columba livia) serving as primary host (BonDurant and Honigberg 1994). In North America, it is speculated that T. gallinae generally occurs wherever domestic pigeons and mourning doves (Zenaida macroura) are found (Stabler 1954, Cole 1999b). This parasite also infects other columbiforms, as well as falcons and hawks, owls, passerine birds, upland birds, waterfowl, and gulls (BonDurant and Honigberg 1994, Cole 1999b).
life cycle and variations Trichomo- nads have the simplest kind of protozoan life cycle, in which the organism occurs only as a trophozoite, with no cyst. These parasites multiply by binary fission (Leber and Novak 1999).
reservoirs and transmission The common pigeon (Columba livia) is believed to be the ultimate source of T. gallinae infection throughout the world (BonDurant and Honigberg 1994). The parasite was introduced to the United States with the pigeons and doves brought by European settlers (Stabler 1954).
Most nestlings become infected and, once acquired, the infections can last up to 2 years (BonDurant and Honigberg 1994). Even though individual birds can lose an infection, the pattern of communal living helps ensure that most birds remain infected (BonDurant and Honigberg 1994).Because there is no resistant cyst stage, transmission from host to host must be relatively direct. In pigeons, the parasite is spread during courtship behavior and infection of young through the process of regurgitive feeding of partially digested foodstuffs (“pigeon's milk”) produced in the crops of the adults (BonDurant and Honigberg 1994). Because T. gallinae is not found in the intestinal tract past the muscular stomach, it cannot be transmitted in fecal droppings; rather, it is transmitted only from the mouth, and secondarily the nares and eyes (BonDurant and Honigberg 1994). Transmission to species other than pigeons is through ingestion of parasites from water contaminated by infected pigeons, or through ingestion of infected hosts (Stabler 1954, Pokras et al. 1993, Cole 1999b).
clinical effects and diagnosis Although trichomoniasis is a general term for an infection, the disease also is termed “canker” in columbiform birds and “frounce” in raptors (Pokras et al. 1993). For columbiforms, most T. gallinae strains are nonpathogenic or only moderately pathogenic for their avian hosts; however, there also are a number of virulent strains (BonDurant and Honigberg 1994). Virulent strains kill young birds, and thus often are selected out of a population. Benign strains may provide immunity against the more virulent strains.
The normal sites of infections in colum- biforms are the mouth, pharynx, esophagus, and crop. Virulent strains may cause lesions in the upper digestive tract and spread to other parts of the body through these ulcerations (BonDurant and Honigberg 1994). Infected birds often appear listless, with ruffled feathers, and with yellowish caseous lesions around the beak or eyes.
Wet, sticky discharges and nodules within the mouth are characteristic of acute disease, whereas hard caseous lesions are associated with chronic infections (Cole 1999b). Lesions in the pharynx or crop may spread to the esophagus and eventually block it, resulting in starvation or even suffocation (Cole 1999b).Trichomonas gallinae can be diagnosed in wet smears from the mouth and oropharyngeal cavity of pigeons or by cultivation of swabs taken from those areas (BonDurant and Honigberg 1994). Stabler argued that examining samples directly in saline solution was better than trying to culture organisms. He described the following technique:
Sit, place the pigeon on its back on the examiner’s left leg; place the floor of the thumb against the tip of the lower mandible and pull back gently, thus opening the bird’s mouth; with a pair of curved forceps, stroke the roof of the mouth back of the palatal flaps in an effort to remove saliva for examination (the curve ofthe forceps should stroke and lift the rear of the flaps); dab saliva in a drop of saline and examine on ’scope. This procedure, when mastered, is quick, easy, and much more efficient than all the swabbing and culturing! (Stabler 1975)
We found this technique to work very well. More recently, the InPouch TF culture system (BioMed Diagnostics, San Jose, California) has been very effective compared with other techniques, including wet mount and direct microscopic examinations, for detecting T. gallinae (Cover et al. 1994, Bunbury et al. 2005).
population effects Trichomonas gallinae is a very common parasite of pigeons and sometimes can cause mortality involving thousands to tens of thousands of birds (Cole 1999b, Hofle et al. 2004). Trichomoniasis is considered by many avian disease specialists to be the most important disease of mourning doves (Zenaida macroura) in North America (Conti 1993). Band-tailed pigeons (Columbafasciata) also have experienced considerable losses (Cole 1999b). The role of T.
gallinae as an agent causing declines or suppression of wild populations is not clear, but in one study on mourning doves T. gallinae was not a significant contributor to a population decline (Ostrand et al. 1995).Trichomoniasis in raptors (frounce) has caused mortality in falcons, and less commonly in hawks and owls, for hundreds of years (Bert 1619, Stabler 1954, Cooper 2002), and may have serious impacts on some raptors populations (Cooper and Petty 1988, Pepler and Oettle 1992). Trichomoniasis among wild raptors is linked to their feeding on urban pigeons. With Cooper’s hawks (Accipiter cooperii), prevalence of T. gallinae was significantly higher among urban nestlings (85%) than nestlings in undeveloped natural areas (exurban sites) (9%) (Boal et al. 1998); nestling mortality also was higher in urban than in exurban nests (Boal and Mannan 1999). In contrast, very few adults were found infected (Boal et al. 1998). In a study among northern goshawks (Accipiter gentilis), overall nestling prevalence among urban birds was 65% and tended to increase with age of nestlings and brood size (Krone et al. 2005). However, prevalences of T. gallinae in raptors can vary considerably by site (Rosenfield et al. 2002), and the overall impact of this parasite on raptors is not well understood.
special problems Although the parasite has a very broad distribution, introduction of T. gallinae to new sites has been of some concern. Domestic pigeons were introduced to the Galapagos Islands in 1972 or 1973 (Harmon et al. 1987). A high prevalence later was found among these introduced birds, along with T. gallinae infections among endemic Galapagos doves (Zenaida galapagoensis); doves examined on pigeon-free islands were not infected (Harmon et al. 1987).
control For virulent T. gallinae strains, removing infected individuals and any other source of infection is an effective strategy to protect susceptible birds (BonDurant and Honigberg 1994). Preventing concentrations of doves at bird feeders and artificial watering sites can help reduce transmission among wild populations (Cole 1999b).
Although environmental persistence of T. gallinae is of short duration, bird feeders and other sites of columbiform concentrations should be kept clean and fresh (Cole 1999b).Most common antibiotics are ineffective against T. gallinae infections and are not practical for wild populations (BonDurant and Honigberg 1994, Cole 1999b). Several nitroimidazole compounds are effective when administered in drinking water, but this group of drugs cannot be used for any food-producing animals in the United States (McDougald 1991, BonDurant and Honigberg 1994).
Cyst Stage Present
EIMERIA spp.
causative agent (classification, morphology) Members of the genus Eimeria fall within the Supergroup Chromoalveolata (App. ι: Table 5) (Adl et al. 2005). As Apicom- plexa, all are parasitic (Lindsay and Todd 1993). Most Eimeria spp. are parasites of the gastrointestinal tract, although some also invade other tissues (Lindsay and Todd 1993).
host range and distribution Members of the genus Eimeria have a worldwide distribution, with many species affecting a wide variety of hosts among all classes of vertebrates (Gylstorff and Grimm 1987, Cole 1999a, Friend and Franson 1999, Bush et al. 2001, Duszynski and Upton 2001). Well over 800 species of Eimeria are reported from mammals alone, and this may be only a fraction of the total in mammals (Duszynski and Upton 2001).
While at least 60 avian species of Eimeria have been reported (Todd and Hammond 1971), many of these species are not well understood. Much of the information on avian coccidiosis is through work on domestic birds. There currently are seven recognized species of Eimeria in chickens, at least four species in domestic turkeys, 10 species in pheasants, two species in guinea fowl, at least six from partridge and quail, and about 15 species in geese (Long 1993). At least six species have been described from raptors (Cawthorn 1993).
life cycle and variations All members of the genus Eimeria have a direct life cycle (Fig.
6.3) that commonly requires about a 1-week period. Eimeria spp. sporozoites infect cells of the intestinal mucosa and may cause serious disease in some hosts. Thick-walled and environmentally resistant oocysts are produced and excreted into the environment. Each mature oocysts contains four sporocysts; each sporocyst has two sporozoites (Bush et al. 2001). Sporogony requires moisture and oxygen, and is most effective between 10 and 35°C (Lindsay and Todd 1993).reservoirs and transmission It is not clear whether most species of Eimeria have only a single vertebrate host (Lindsay and Todd 1993, Bush et al. 2001) or if they regularly can infect more than one species (Duszynski and Upton 2001). Among wild mammals, members of the Orders Rodentia and Artiodactyla have the largest numbers of known Eimeria species, followed by Lagomorpha, Carnivora, Diprodon- tia, Insectivora, and Chiroptera (Duszynski and Upton 2001). Eimeria spp. infections are reported among wild birds of the Orders Anseriformes, Galliformes, and Passeriformes (Todd and Hammond 1971). Transmission typically is by ingestion of a mature oocyst containing infective sporozoites (Duszynski and Upton 2001).
clinical effects and diagnosis Enteritis, associated with diarrhea, is a common clinical syndrome. Some Eimeria spp. also may invade the liver, leading to hepatitis. However, pathogenicity of coccidian parasites in wild mammals may not be of great significance, and appears to be a disease due primarily to human intervention (Duszynski and Upton 2001).
Among geese, there is a renal and an intestinal form of coccidiosis (Long 1993). Renal coc- cidiosis, caused by E. truncata, develops only in the kidneys and the cloaca near its junction with the ureters; the route to infecting the kidneys is unknown (Long 1993). Most other avian species of Eimeria are not considered severely pathogenic (Long 1993).
When a host is not killed, diagnosis of a coc- cidian infection typically depends on finding oocysts in the feces and observing them completely sporulate (Duszynski and Upton 2001). The polymerase chain reaction and random amplified polymorphic DNA tests also are used to detect and identify coccidia (Comes et al. 1996, Barta et al. 1997, Duszynski and Upton 2001).
population effects Among mammals, Eimeria stiedae, for example, may be a limiting factor on young wild rabbits (Oryctolagus cuniculus) of Australia in normal and wet years, but not in dry years (Dunsmore 1971). Most wild birds pass small numbers of oocysts in their feces without apparent ill effects, but coccidiosis becomes important as a disease when animals live under crowded or dense populations that allow buildup of infective oocysts in the environment and ingestion by susceptible hosts (Long 1993). Among domestic birds, it is speculated that the poultry industry is dependent on continuous medication in the feed (Long 1993).
Renal coccidiosis, caused by E. truncata, has caused losses as high as 87% among infected, presumably domestic, flocks of birds in Iowa (Long 1993). This parasite also has been associated with mortality among Canada geese in North America (Critcher 1950, Farr 1954, Hanson et al. 1957)
special problems No ongoing special problems are noted for Eimeria spp.
control Among domestic birds, there are a wide variety of anti-coccidial chemotherapies used to combat Eimeria infections (Long 1993). There also have been some efforts to immunize domestic birds by use of attenuated strains of Eimeria, as well as vaccinating with specific antigens (Long 1993). Immunity does not play a significant role; among domestic birds immunity lasts only 4 to 6 weeks in birds recovering from infection (Long 1993).
Indirect (Heteroxenous) Life Cycles
Use Vertebrates as Both Intermediate and Definitive Hosts
TOXOPLASMA CONDll
CAUSATIVE agent (CLASSIFICATION, morphology) Toxoplasma gondii is classified as a coccidian parasite, with Eimeria and Sarcocystis, among the Apicomplexa in the Chromoalveolata (Dubey and Odening 2001). The name Toxoplasma (toxon: arc, plasma: form) is derived from the crescent shape of the tachyzoite stage (Hill et al. 2005). Toxoplasma gondii is the sole member of the genus (Hill et al. 2005); currently, three dominant genotypes (I, II, III) are recognized in the species (Grigg et al. 2001, Su et al. 2003, Volkman and Hartl 2003). Recently, another type (X) has been observed in marine mammals (Miller et al. 2004).
host range and distribution Although it can only complete its life cycle in felids (Dubey and Odening 2001), T. gondii can infect nearly all mammal and bird species and has a worldwide distribution (Lehmann et al. 2006). In recent years, it has been described and become widely reported among marine mammals (Dubey 2004). Toxoplasma gondii is postulated to originally have been a parasite of South American felids that underwent a relatively recent worldwide expansion from the effects of both migratory birds and the expansion of maritime travel; in particular, the transatlantic slave trade may have played a considerable role in this expansion (Lehmann et al. 2006).
life cycle and variations Wild and domestic felids are the only known definitive hosts; oocysts that successfully sporulate in the environment and infect susceptible hosts are formed only in cats (Fig. 6.9: (Dubey
et al. 1970). In contrast to other coccidia, several life history stages of T. gondii can infect both intermediate hosts and definitive hosts. The three infectious stages of T. gondii are sporozoites, tachyzoites (trophozoites), and brady- zoites, developing in meront-like tissue cysts called pseudocysts) (Dubey 2004).
Most natural infections probably are acquired by ingesting sporulated oocysts with sporozoites in food or water contaminated with cat feces, or by ingesting pseudocysts in infected meat (Hill et al. 2005). Oocysts transform into tachyzoites shortly after ingestion by intermediate hosts (Fig. 6.9: Γa). Tachyzoites rapidly proliferate in pseudocysts by binary fission (Bush et al. 2001) and destroy infected cells during acute infection. Tachyzoites localize in neural and muscle tissue and develop into bradyzoites within pseudocysts (Fig. 6.9: Γ^). Bradyzoites multiply slowly in tissue cysts (Bush et al. 2001). Cats become infected by consuming intermediate hosts with these tissue cysts (Fig. 6.9: ι,Λ).
Toxoplasma gondii also may spread by infected macrophages to mesenteric lymph nodes and then to distant organs by invasion of the lymphatic system and blood (Hill et al. 2005). The host cells die and release tachyzoites, which invade adjacent cells and continue the process (Wilson and McAuley 1999, Hill et al. 2005). Tachyzoites are pressured by host immune responses to transform into bradyzoites and to form meront-like pseudocysts (Wilson and McAuley 1999). Although
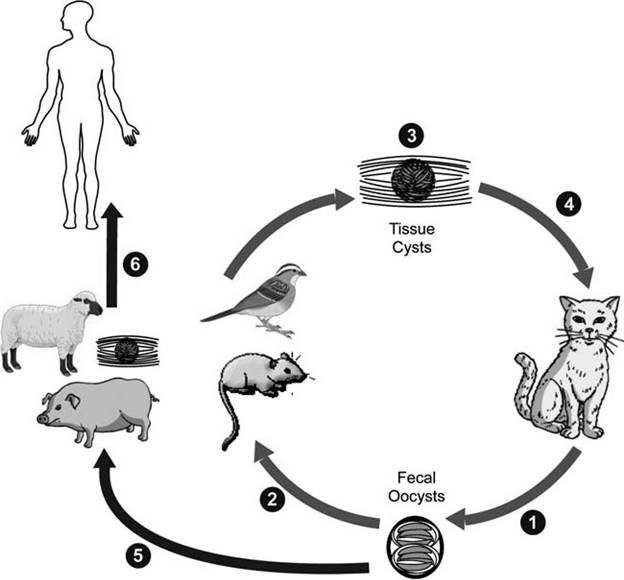
FIGURE 6.9 Life cycle of Toxoplasmagondii. Three sources of infection (a, b, c) occur for both intermediate and definitive hosts. Bradyzoites in pseudocysts will become tachyzoites if ingested by a new host, and then form bradyzoites in that intermediate host; or they will go on to form gametes and oocysts if they are in a felid definitive host (Courtesy of Centers for Disease Control and Prevention's Division of Parasitic Diseases and Malaria, www.dpd.cdc.gov/dpdx).
pseudocysts containing bradyzoites may develop in visceral organs, including lungs, liver, and kidneys, they are more prevalent in muscular and neural tissues, including the brain, eye, skeletal, and cardiac muscle (Hill et al. 2005). Infected intermediate hosts may contain infective cysts for their lifetime and shed tachyzo- ites, but they cannot complete the life cycle (Hill et al. 2005).
Generally, cats are more readily infected by ingesting bradyzoites than oocysts (Dubey 2004); ingesting as few as one bradyzoite can lead to a cat shedding millions of oocysts (Dubey 2001). Virtually all cats ingesting pseudocysts containing bradyzoites will shed oocysts, whereas less than half of the cats ingesting tachyzoites or oocysts will complete the life cycle (Dubey and Frenkel 1972).
When felids ingest pseudocysts from infected prey, the released bradyzoites penetrate epithelial cells of the small intestine (lamina propria) (Dubey 2004). In the epithelial cells, T. gondii multiplies (enteroepithelial cycle) and forms meronts (also called schizonts), which, in turn, continue developing to merozoites, gametes, zygotes, and oocysts (Dubey and Frenkel 1972, Hill et al. 2005). Formed oocysts are discharged into the intestinal tract by rupture of the epithelial cells and sporulate about 1-5 days after passing into the environment (Hill et al. 2005).
As this enteroepithelial cycle progresses, bradyzoites also penetrate the feline intestine and multiply as tachyzoites, eventually disseminating to other tissues in the cat. Both intestinal and extraintestinal stages of T. gondii may occur for several months, or even remain for the life time of the cat (Hill et al. 2005).
Often, wild or domestic animals can become infected (Fig. 6.9: ι^). Humans become infected by eating these animals, as well as by eating food or water contaminated by cat feces, including from cat litter boxes (Fig. 6.9: l6). In a more serious problem, developing fetuses may be infected transplacentally, sometimes with fetal mortality or severe neurological consequences after birth.
RESERVOIRS AND TRANSMISSION One collective population estimate of domestic and feral cats in the United States ranges from about 93 to 133 million cats [Introduced Species Summary Project: Domestic Cat (Felis catus), www.columbia.edu/itc/cerc/danoff-burg/invasion_bio /inv_spp_summ/Felis_catus.html]. Up to 2% of all cats are estimated to be shedding T. gondii oocysts at any given time, with occasional higher prevalences reported (Dubey 2004).
Three known routes of transmission for T. gondii include transmission through oocysts shed in the feces of wild and domestic felids, ingestion of tissue cysts from meat, and congenital transmission (Fayer et al. 2004). The oocysts are the only stage to enter the general environment, and they can remain infectious for months to years under moist temperate conditions while being widely dispersed in large numbers (Yilmaz and Hopkins 1972, Frenkel et al. 1975); thus, oocysts are viewed as the most likely means of transmission to non-carnivores (Fayer et al. 2004). Land-based surface runoff is a likely source of T. gondii oocysts for marine mammals (Miller et al. 2002a).
Immunity does not eradicate infection, and T. gondii tissue cysts persist several years after acute infections (Hill et al. 2005). It is not clear how T. gondii is destroyed in immune cells (Renold et al. 1992). Extracellular forms of the parasite are directly affected by antibody, but intracellular forms are not; it is believed that T-lymphocytes and lymphokines are more important than humoral factors in immune destruction of T. gondii (Renold et al. 1992).
clinical effects and diagnosis Pathogenicity of T. gondii is determined by many factors, including the susceptibility of the host species, virulence of the parasitic strain, and the life cycle stage ingested; oocyst-induced infections are the most severe clinically in intermediate hosts, independent of dose (Dubey 2004). Although cats of any age can die of toxoplasmosis, kittens and those with depressed immunity are the most susceptible (Dubey and Carpenter 1993a, 1993b). Toxoplasmosis can be a severe disease in domestic animals, and may cause embryonic and fetal death, abortion, stillbirth, and neonatal death (Dubey 2004).
Clinical signs of toxoplasmosis are nonspecific and mimic those of several other diseases (Dubey and Odening 2001). Trophozoites can invade many tissues, leading to fever, splenomegaly, myocarditis, pneumonitis, and encephalitis. The pathology described among marine mammals includes meningoencephalitis, with lesser observations of hepatitis, myocarditis, and lymphadenitis, as well as disseminated and congenital infections (Dubey et al. 2003, Kreuder et al. 2003, Dubey et al. 2004, Miller et al. 2004).
Diagnosis of toxoplasmosis is made by biologic, serologic, or histologic methods, or by their combination (Hill et al. 2005). Oocysts in infected cats are diagnosed by concentration methods such as flotation in high-density sucrose solutions (Ruiz and Frenkel 1980). Isolation of T. gondii from animals by inoculation of laboratory animals and tissue cultures can lead to definitive diagnoses (Dubey and Odening 2001). However, for epizootiological surveys, serological prevalence is a better measure of T. gondii prevalence (Dubey 2004). Serological tests include the dye test, indirect fluorescent antibody test (Miller et al. 2002b), modified agglutination test (MAT), ELISA, and immunosorbent agglutination assay test (Dubey and Beattie 1988). Some work with polymerase chain reaction is promising (Hill et al. 2005).
population effects Generally, the parasite causes few problems among wildlife populations. One exception may be with sea otters (Enhydra lutris), and this is addressed in the next section.
special problems Toxoplasma gondii has been detected in a wide variety of geographic areas from seals (Lambourn et al. 2001, Dubey et al. 2003, Measures et al. 2004, Honnold et al. 2005), dolphins (Dubey et al. 2003, Cabezon et al. 2004, Dubey et al. 2005), sea otters (Cole et al. 2000, Lindsay et al. 2001b, Miller et al. 2002b, Dubey et al. 2003), and other marine mammals (Cruikshank et al. 1990, Dubey et al. 2003, Philippa et al. 2004). Pathology and mortality have been described from dolphins (Inskeep et al. 1990, Jardine and Dubey 2002, Resendes et al. 2002), a seal (Dubey et al. 2004), and sea otters (Kreuder et al. 2003, Miller et al. 2004). In an experimental infection, Toxoplasma established patent infections in gray seals (Halichoerus grypus), but did not cause pathology (Gajadhar et al. 2004).
The southern sea otter (Enhydra lutris nereis) is listed as threatened under the Endangered Species Act and is considered a keystone species strongly influencing the abundance and diversity of other species within the kelp forest ecosystem (Jessup et al. 2004). There has been a recent decline as a consequence of high mortality rather than low recruitment, with considerable concern among biologists and managers (Estes et al. 2003, Gerber et al. 2004). Current evidence is that toxoplasmosis is making a significant contribution to this mortality (Kreuder et al. 2003, Hill et al. 2005). Of the T. gondii evaluated from sea otters in one study, 40% were Type II and 60% were Type X; no Types I or III were observed (Miller et al. 2004). Subsequent work has an estimate of 72% Type X in sea otters, with the observation that the same Type X has been isolated from other marine mammals as well (Conrad et al. 2005).
Of the potential sources of infection, ingestion of sporulated oocysts seems most likely; sea otters do not prey on warm-blooded animals, which are the usual intermediate hosts of T. gondii (Conrad et al. 2005). There is evidence that Toxoplasma gondii has invaded the marine ecosystem through runoff from terrestrial sources (Miller et al. 2002a, Fayer et al. 2004). Sea otters in areas of high freshwater runoff are more likely to be exposed to T. gondii than otters from low or medium freshwater runoff (Miller et al. 2002a). Increased risk of sea otter mortality has been linked to increased exposure to T. gondii (Miller et al. 2002a, Kreuder et al. 2003, Conrad et al. 2005).
Collectively, there is good evidence that Toxoplasma oocysts are spread from terrestrial runoff into the marine ecosystems, where they are concentrated in the resident shellfish; shellfish are a major food source of the California sea otters and serve as a means of transmission (Mitchell and Sinai 2006), or as paratenic hosts (Arkush et al. 2003). Eastern oysters (Crassostrea virginica) are able to remove T. gondii oocysts from seawater readily and retain their infectivity for at least 85 days (Lindsay et al. 2001a, 2004). Similarly, another bivalve mussel, Mytilus galloprovincialis, also is able to take up and retain infective T. gondii oocysts (Arkush et al. 2003). It also has been observed that anchovies (Family Engraulidae), a small common saltwater fish, can be infected with T. gondii after exposure (Massie and Black 2008); these fish are common prey items of marine mammals. Thus far, natural infections have not been reported in these species, however, and their actual role in the epizootiology of the disease is unclear (Conrad et al. 2005). Interestingly, the oocysts also sporu- late readily and remain viable in seawater for at least 6 months (Lindsay et al. 2003).
Reasons for the unusual morbidity and mortality from toxoplasmosis in sea otters, compared to the subclinical or mild infections seen in most immunocompetent terrestrial animals, are unclear (Conrad et al. 2005).
control Prevention of Toxoplasma among humans and domestic animals generally is by maintaining cleanliness and reducing ingestion of contaminated food or water (Hill et al. 2005). There are no specific recommendations to prevent transmission of T. gondii by drinking water because the level of contamination is unknown and because it can be transmitted by several modes (Dubey 2004). At present there is no vaccine to control toxoplasmosis in any species (Dubey and Odening 2001).
Arthropod-borne Apicomplexa
PLASMODIUM RELICTUM (AVIAN MALARIA)
causative agent As Apicomplexa (Adl et al. 2005), all members of the genus Plasmodium are parasitic. There are about 34 recognized species of Plasmodium described from birds (Bennett et al. 1993). Over 25% of all avian host species are parasitized by members of the genus Plasmodium (Bennett et al. 1982, 1994).
host range and distribution Plasmodium relictum has a worldwide distribution (Bennett et al. 1993, Peirce 2005, Kimura et al. 2006); it is the most common avian Plasmodium species and has been reported from over 300 species and at least 70 families of birds (Bennett et al. 1993, Valkiunas 2005).
life cycle and variations Using the general malarial life cycle model (Fig. 6.5), P. relictum sporozoites are introduced to susceptible birds by the bites of infected culicine mosquitoes. Sporozoites invade a variety of cells, including the Malpighian body, and later the lungs, brain, and spleen (van Riper et al. 1994). After three or more generations of merogony in these tissues, the merozoites invade erythrocytes, where they also undergo merogony; in older literature this was referred to as exoeryth- rocytic (out of erythrocyte) schizogony and erythrocytic schizogony. The resulting merozo- ites then develop into gametocytes (van Riper et al. 1994).
Gametocytes ingested by Culex spp. mosquitoes form gametes that fuse into zygotes that, in turn, become motile (ookinetes); ookinetes penetrate midgut epithelial cells and form an oocyst on the hemocoel side of the gut. Following sporogony, oocysts rupture and sporozoites are released into the hemocoel of the mosquito; from here they migrate to the salivary glands to be injected into the new avian host during a blood meal by the mosquito (van Riper et al. 1994).
reservoirs and transmission The avian malaria cycle requires the avian vertebrate host and culicine mosquitoes as the invertebrate hosts (van Riper et al. 1994). Although P. relictum completes its development in at least 26 species of mosquitoes (Huff 1965, Garnham 1966), culicine mosquitoes are judged to be the most likely transmitters under natural conditions (van Riper et al. 1994).
clinical effects and diagnosis Clinically ill birds may appear listless, have ruffled feathers, lose their appetite, and have difficulty breathing; pathologic changes often involve anemia as well as enlargement of the spleen and liver (van Riper et al. 1994). Some infected birds may show few if any symptoms.
population effects The parasite P. relictum Capistranoae has been associated with severe population declines of native birds in Hawaii, including extinction of many native species of Hawaiian honeycreepers (Drepanidinae). These events followed introduction of the vector, the house mosquito, Culex quinquefasciatus, in approximately 1826 and introduction of the parasite through migratory birds (Warner 1968, van Riper et al. 1986).
special problems Endemic Hawaiian forest birds are highly susceptible to infection with Plasmodium relictum capistranoae, with some having mortality rates of 65-90% after being bitten by a single infective mosquito (Atkinson et al. 1995, 2000, 2001b). High susceptibility of native Hawaiian honeycreepers (Family Fringillidae, Subfamily Drepanid- inae) to avian malaria is believed to be one of the primary factors responsible for the disappearance of these birds from lowland habitats on the main Hawaiian islands (Atkinson et al. 2001a). There is compelling evidence that the near absence of native forest birds below elevations of 900 m in Hawaii was due to the Culex spp. vector and parasite transmission (van Riper et al. 1986). With further work, it was established that malaria transmission was greatest in mid-elevation forests where the highly susceptible native forest birds overlapped in distribution with mosquitoes and where these native birds themselves served as the primary reservoirs of infection (van Riper et al. 1986, Atkinson et al. 2001a, Woodworth et al. 2005). The native birds were almost entirely absent from low-elevation study areas because of high mosquito densities; malaria prevalence in the primarily exotic, diseaseresistant lowland avifauna was low (van Riper et al. 1986, Woodworth et al. 2005).
There is evidence that at least one native Hawaiian forest bird, the amakihi (Hemignathus virens), has endured the parasite, has mounted an immune response (Atkinson et al. 2001a), and is even recolonizing low-elevation habitats that contain higher densities of the culicine vector (Woodworth et al. 2005).
control No specific measures are taken to control avian malaria in wild birds.
TRYPANOSOMA SPP. (NAGANA)
CAUSATIVE AGENT (CLASSIFICATION, morphology) The nagana disease complex involves three species of trypanosomes (T. vivax, T. congolense, T. brucei brucei) (Logan-Henfrey et al. 1992); they are classified in the Kinetoplastea among the Euglenozoa in the Excavata Supergroup (Adl et al. 2005). These are long, spindleshaped cells (about 2 ? 20 mμ) with a single flagellum (Logan-Henfrey et al. 1992). All three are salivarian parasites typically passed through the bites of tsetse flies.
host range and distribution Distribution of this group of parasites is between 14°N and 29°S on the African continent (Wells and Lumsden 1969). Distribution is closely linked to the distribution of tsetse flies (Glossina spp.), which infest about 11 million km2 of Africa, or about one-half of the available arable land. A wide variety of African ungulates are infected with this complex of organisms (Ashcroft 1959, Baker et al. 1967, Geigy et al. 1967, Baker 1969). Most of these wild ungulates are resistant to the trypanosomes, but occasional pathology and disease has been reported among them (Molyneux 1982).
LIFE cycle and variations The trypomas- tigote and the epimastigote (Fig. 6.8) stages are found among hosts of the African salivar- ian trypanosomes (Logan-Henfrey et al. 1992). Infected tsetse flies inoculate metacyclic trypo- mastigotes into the skin of animals, where the trypanosomes grow for several days and cause local swellings. They enter the lymph nodes and then the bloodstream, where they divide rapidly by binary fission (Fraser and Mays 1986).
After the tsetse fly feeds on a host infected with trypanosomes, the bloodstream trypomas- tigotes develop into procyclic trypomastigotes in the midgut, then into epimastigotes, and then into metacyclic trypomastigotes (Logan- Henfrey et al. 1992). There also are a number of variations in the timing of the life cycle, the various stages developed in the hosts, and the host sites at which the parasites concentrate among the trypanosome species (Fig. 6.10) (Logan-Henfrey et al. 1992).
reservoirs and transmission These African trypanosomes typically are passed by the bite of an infected tsetse fly. There are potentially 23 tsetse fly species involved, classified into three taxonomic groups according to their preferred ecological habitat. The Glossina morsitans group typically is found in savanna, the G. palpalis group prefers areas around rivers and lakes, and the G. fusca group lives in high forest areas (Logan-Henfrey et al. 1992). The limits of tsetse infestation are determined primarily by climate and secondarily by vegetation (Ford 1971, Logan-Henfrey et al. 1992).
In addition to the typical transmission by hematophagous arthropods, there also is evidence that transmission of trypanosomes to predators such as lions (Panthera leo) and hyenas (Crocuta crocuta) can occur by ingestion of infected ungulates (Baker et al. 1967, Baker 1969, Geigy and Kauffman 1973). This insight is supported by laboratory studies (Heisch 1963, Moloo et al. 1973).
CLINICAL EFFECTS AND DIAGNOSIS The severity of disease varies with the species of animal and the trypanosome species involved (Molyneux 1982). There are individual case reports of wild ungulate species in which lesions were attributed to trypanosome infections, but generally African ungulates have an innate tolerance of trypanosome infections (Molyneux 1982). For cattle and other susceptible species, the primary clinical sign is anemia, with an associated stunting and wasting
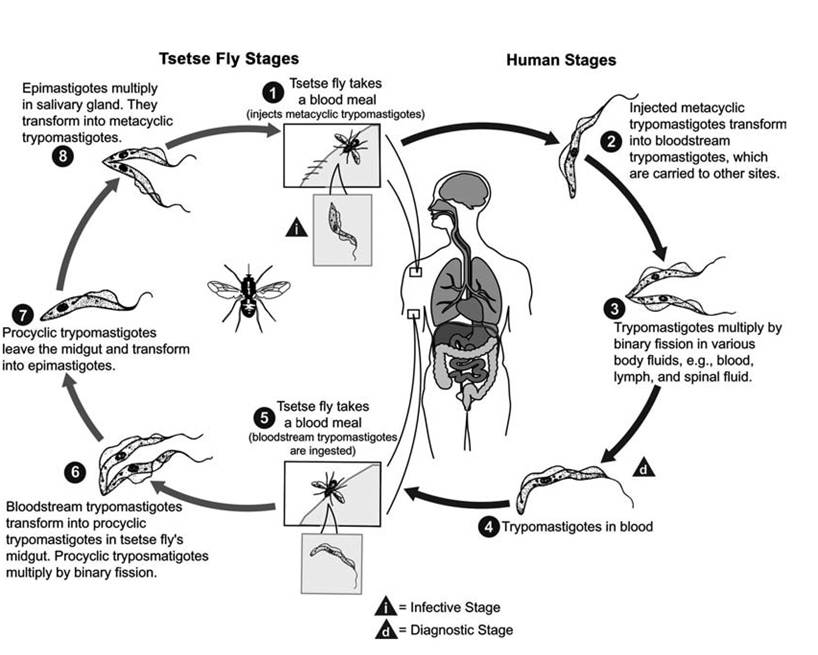
FIGURE 6.10 Life Cycle of a Salivaria (Trypanosoma brucei gambiense) (Courtesy of Centers for Disease Control and Prevention’s Division of Parasitic Diseases and Malaria, www.dpd.cdc.gov/dpdx).
(Molyneux 1982). Most infected cattle have a chronic disease course with high mortality (Fraser and Mays 1986); mortality usually is associated with circulatory disorder, damage to the heart, and the anemia (Molyneux 1982). There are few consistent clinical signs or internal lesions specific to the disease, and other diseases in Africa (e.g., anaplasmosis, babesiosis, theileriosis) can be confused with trypanosomiasis (Logan-Henfrey et al. 1992).
Trypanosome infections may be detected directly by microscopic observation of the parasites, by immunoassays for trypanosome antigens, or by biochemical techniques that detect trypanosomal DNA. Identifying trypanosome infections also is based on serological detection of anti-trypanosomal antibodies (Logan- Henfrey et al. 1992).
population effects There is little overall impact on indigenous wild ungulate species. However, much of Africa between 14°N and 29°S was excluded for cattle due to the severe impacts of trypanosomiasis. There is virtually no overlap between the cattle-raising regions of Africa and the range of tsetse flies and the trypanosomes (Donelson and Turner 1985).
special problems Cattle have been denied vast areas of Africa by the occurrence of trypanosomiasis. Trypanosomiasis is regarded economically as the most important livestock disease in Africa (Jawara 1990), with an estimated annual loss to African farmers in meat production alone estimated at US$5 billion in the 1990s (Gyening 1990, Logan-Henfrey et al. 1992).
control Historically, risk of trypanosomiasis was minimized by avoiding tsetse-infected areas. More recently, control of trypanosomiasis in livestock relies on the treatment of the infected host and on vector control (Logan- Henfrey et al. 1992).
In the early 1900s, it was observed that the mortality among wild ruminants from rinderpest was so high that the tsetse fly, especially Glossina morsitans, was severely reduced in some areas such as Kruger National Park, from lack of suitable hosts (Stevenson-Hamilton 1911, Duke 1919, Carmichael 1933). At that time, “game elimination” became a technique used to limit the sources of blood meals available to the tsetse flies, and to eliminate trypanosome reservoirs (Logan-Henfrey et al. 1992); this in turn was intended to reduce tsetse fly populations, specifically Glossina morsitans, and to enhanced success of cattle in the region; there was some evidence for success with this approach (Clarke 1964, Ford 1971). Between the 1920s and the 1960s, it was estimated that 1.3 million game animals were killed in tsetse control programs (Matthiessen and Douthwaite 1985). However, the public outcry against wildlife hunting as a means of tsetse control (Cockbill 1967) and the developing evidence for its overall inefficiency (Glover 1965), along with the advent of insecticides, largely ended the practice by the early 1970s (Matthiessen and Douthwaite 1985). At that time there also was an increasing appreciation of the game animals as valuable in themselves and as important national resources (Molyneux 1982).
Vector control historically involved bush clearing in an effort to destroy tsetse fly habitat, to limit the sources of blood meals available to the tsetse flies, and to eliminate trypanosome reservoirs (Logan-Henfrey et al. 1992). Pastoralists and arable farmers tend to clear brush as part of their land use, making the land unsuitable for the flies (Matthiessen and Douthwaite 1985). Some savanna woodland clearing was done to destroy G. morsitans; while there is evidence that this has been effective in some regions, there is no overall assessment of its effects, but there is speculation that continued deforestation and other habitat changes changed and reduced the occurrence of tsetse flies, and trypanosomiasis, in parts of Africa (Molyneux 1982).
Game destruction and bush clearing were replaced by insecticides for tsetse control (Jordan 1974, Allsopp 1984). A variety of insecticides have been used, but generally have had limited effectiveness and often have had significant side effects on nontarget species (Du Toit 1954, Graham 1964, Wilson 1972, Molyneux 1982, Matthiessen 1985, Matthiessen and Douthwaite 1985).
In recent years, use of traps and screens impregnated with pyrethrum has been effective in reducing tsetse flies of the G. palpalis and
G. fusca groups; also, a number of attractants are effective for capture of the G. morsitans group of flies (Logan-Henfrey et al. 1992). There has been considerable interest in biological control, including use of sterilized males, but no effective techniques have emerged (Logan-Henfrey et al. 1992). There also is exploration of developing more trypanotoler- ant animals, but these animals represent only a small portion of the cattle in Africa (Logan- Henfrey et al. 1992).
Treatment of individually affected animals can be accomplished with a number of drugs, including dimazene aceturate, quinapuramine sulfate, and homidium bromide, among others (Logan-Henfrey et al. 1992). There is little likelihood of an effective vaccine because of the unique ability of trypanosomes to evade the immune system while in the blood stream by switching on new genes to encode new surface antigens (Donelson and Turner 1985). Trypano - somes may possess up to 1,000 variable surface glycoprotein (VSG) genes and, from these, they select certain ones to express when they are at the metacyclic stage of development, others as trypomastigotes in the early stages of infection in the blood, and still others during the later stages of chronic (Boothroyd 1985, Borst 1986, Van der Ploeg 1987, Pays and Steinert 1988).
Helminth-borne
H∣sτoMθNAs Meleacridis
CAUSATIVE AGENT (CLASSIFICATION, morphology) Histomonas meleagridis is included among the Trichomonadida of the Parabasalia, in the Supergroup Excavata (Adl et al. 2005). Histomonas meleagridis is one of two species in this genus and the only one of pathogenic significance. Two forms of H. meleagridis have been described, a lumen form with one or two flagellae, and a tissue form without flagellae; both range from 6 to 20 mμ in size (McDougald and Reid 1978). There is also some evidence for a third form parasitizing gonads of larvae of the cecal nematode Heterakis gallinae; a 3 μm form has been observed in cecal worm eggs (BonDurant and Wakenell 1994).
cost RANGE AND DISTRIBUTION Most, if not all, gallinaceous birds are susceptible hosts (Cole and Friend 1999), including chukar (Alectoris chukar), pheasant (Phasianus colchicus), quail (Order Galliformes), grouse (Family Phasianidae), wild turkeys (Meleagris gallopavo), jungle fowl (Gallus gallus), guinea fowl (Family Numididae), and peafowl (Pavo spp.) (BonDurant and Wakenell 1994). Histomonas meleagridis essentially has a worldwide distribution (Cole and Friend 1999).
life cycle and variations Histomonads multiply by binary fission. The H. meleagridis life cycle is unique in that it is one of the rare examples of a helminth-borne life history pattern; in addition, earthworms typically are used as paratenic hosts (Cole and Friend 1999). Histomonas meleagridis uses the cecal nematode Heterakis gallinae as a vector for entry into the avian hosts (BonDurant and Wakenell 1994). Although histomonads can be passed directly under optimal circumstances, H. meleagridis quickly loses viability upon cooling outside the host or inside a dead host; thus, development of some resting stage inside the nematode eggs appears essential for survival of H. gallinae, and this nematode is considered critical to the natural transmission of H. meleagridis (BonDurant and Wakenell 1994).
While the histomonads are carried inside the nematode egg (Tyzzer 1934), it still is not clear how the parasite becomes incorporated into the egg and the nature of the form in which it may exist for several years (BonDurant and Wakenell 1994). Both the male and female
H. gallinae become infected with Histomonas; the histomonads then become incorporated into the eggs of the female (Lee 1970, McDougald 1991). There is some evidence that the male Heterakis may become infected first and transfer the infection to the female during copulation (Springer et al. 1969, McDougald 1991). Unembryonated Heterakis eggs are not infective; also, the processes of hatching or physical destruction of the egg seem important in release of infective histomonads (BonDurant and Wakenell 1994).
reservoirs and transmission Heterakis gallinae, the nematode involved, is an ascarid with a direct life cycle, but one that also commonly uses earthworms as paratenic hosts. After the histomonad eggs are shed into the environment, the embryos develop to second stage larvae (L ). On being ingested, the nematode larvae develop to adulthood in the ceca of their avian hosts. The histomonads accompanying the egg also infect the birds and may cause pathology before infecting the new generation of helminth ova produced by infecting H. gallinae.
In addition, earthworms often ingest fecally contaminated soil containing H. gallinae eggs infected with histomonads (Kemp and Franson 1975, BonDurant and Wakenell 1994). Thus earthworms concentrate the nematode eggs and enhance transmission; earthworms also take the eggs to new sites as they migrate in the soil and help preserve the eggs from fungi and predation by other soil invertebrates (Lund et al. 1966). After earthworms ingest Heterakis gallinarum eggs, L larvae hatch out and invade the earthworm tissues; thus the earthworm concentrates the H. gallinarum larvae as well as the histomonads present (Lund et al. 1966). Susceptible birds consuming these earthworms become infected by both the Heterakis gallinarum nematode and Histomonas meleagri- dis. Thus, while earthworms are not required for the nematode or histomonad life cycle, they are considered to play a critical role in enhancing natural transmission of each (BonDurant and Wakenell 1994).
clinical effects and diagnosis The disease histomoniasis also is called blackhead because infections often cause cyanosis, a blue or black appearance on the skin of the head on turkeys and some other birds due to an excess of reduced hemoglobin in the blood (Cole and Friend 1999). There are no clinical signs specific to histomoniasis. Infected wild turkeys often are listless, amd have ruffled feathers and drooped wings. Feces are often sulfur-yellow in color and, combined with other field signs, are highly suggestive of histomoniasis (Cole and Friend 1999). The term enterohepatitis also is applied, reflecting their invasion of the cecum and, in more serious cases, causing multifocal necrosis in the liver (BonDurant and Wakenell 1994)
Turkeys, grouse, and partridge develop severe disease and have mortality rates that can exceed 75% of the birds infected (Cole and Friend 1999). In contrast, histomoniasis is less severe in gray partridge (Perdix perdix) and northern bobwhites (Colinus virginianus) (Cole and Friend 1999).
Pathologic effects of Histomonas appear to involve an interaction with fecal bacteria. Pathologic effects are fully present only with the full complement of fecal bacteria and are reduced in the presence of single species or limited combinations of fecal bacteria (Franker and Doll 1964, Springer et al. 1970). Histomonas does not colonize germ-free animals (Franker and Doll 1964, Kemp 1974).
A presumptive diagnosis can be based on the presence of characteristic gross pathology and history. Laboratory confirmation generally involves microscopic examination for living flagellates with one or two flagellae, an axostyle- like projection, and a 6-20 mμ diameter size; these organisms also can be cultured in Dwyer's medium (BonDurant and Wakenell 1994).
population effects Turkeys are highly susceptible to Histomonas meleagridis infections (his- tomoniasis, blackhead). The parasite can cause catastrophic losses in both wild and domestic turkey populations (Cole and Friend 1999).
special problems One interesting case study involving Histomonas in wildlife occurred in the southeastern United States; it illustrates the complexity of parasitic interactions with their hosts. In the 1960s, there was evidence that wild turkey populations in the southeastern United States were declining; it was proposed that the turkeys were suffering from histomoniasis transmitted from bobwhites (Colinus virginianus) infected with Histomonas-infected Heterakis gallinarum eggs (Kellogg and Reid 1970). But the bobwhites carried Heterakis bonasae rather than H. gallinarum, and only H. gallinarum is known to carry Histomonas (Kellogg and Reid 1970). Thus bobwhites seemed relatively unimportant in contaminating soil with Histomonas-bearing Heterakis eggs (Lund and Chute 1971). However, Indian red jungle fowl (Gallus gallus murghi) previously released in the range of eastern wild turkey (Meleagris gallopavo silvestris) carried Histomonas spp. and were considered a more likely reservoir host for histomoniasis (Kellogg et al. 1972).
control A variety of anthelminthics are used in commercial operations (BonDurant and Wakenell 1994). Because of the high susceptibility of turkeys compared to chickens, wild and domestic turkeys should not have exposure to habitat frequented by chickens; the development in the turkey industry of confinement rearing has reduced exposure to histomonads (BonDurant and Wakenell 1994).
Opportunistic Soil and Water Organisms
ACANTHAMOEBA SPP.
causative agent (classification, morphology) Acanthamoeba is a genus of amebae that generally are free living, but include several species of that can be pathogenic to wildlife, domestic animals, and humans (Visvesvara 1999, John 2001). The genus derives its name from the distinctive feature of its acanthopodia (Gr. acanth: spine or thorn), tapering spike-like pseudopodia, and is classified in the Amoebo- zoa (Adl et al. 2005). Trophozoites average about 24 to 56 μm in length (John 1993). Species often have been named on the basis of variation in cyst morphology among Acanthamoeba (John 1993), but are not easily distinguished (Costas and Griffiths 1986, John 1993).
host range and distribution Acan- thamoeba are distributed worldwide in soil and water (John 1993). Although generally free-living amebae, they are able to cause disease and even death in numerous animals, including humans (John 2001).
LIFE CYCLE AND VARIATIONS The life cycle is simple and includes a motile feeding trophozoite (ameba stage) undergoing binary fission and a resting cyst stage (John 2001).
reservoirs and transmission Under most conditions, Acanthamoeba spp. are free- living phagotrophs in freshwater ponds, rivers, lakes, soil, sewage, and sludge, feeding mainly on bacteria (Ma et al. 1990, Visvesvara and Stehr-Green 1990, John 2001). They occasionally also have been isolated from brackish water and seawater (Sawyer et al. 1976). There is no evidence for an animal reservoir or carrier host for Acanthamoeba (John 1993).
Infections result from environmental exposure; animal-to-animal transmission is unknown (John 2001). Some infections are associated with aquatic exposure (John 2001). In other cases, infections may occur through the lower respiratory tract or through ulcers of the skin or mucosa, or direct contact with the eye (John 1993).
CLINICAL EFFECTS AND DIAGNOSIS Acan- thamoeba infections can involve the central nervous system, eye, and other organs; invasion of the central nervous system appears to be by the circulation, with the initial focus of amebic infection occurring elsewhere in the body (John 2001). Syndromes can include an amebic meningoencephalitis, granulomatous amebic encephalitis, and keratitis (John 1993). Both the trophozoite and cyst stages can occur in tissues of infected animals (John 2001).
Primary amebic meningoencephalitis usually is fatal in humans (John 1993), and generally is diagnosed by microscopic detection of living or stained amebae in cerebrospinal fluid or by culture (John 2001). Granulomatous amebic encephalitis is diagnosed by microscopic detection of trophozoites in cerebrospinal fluid or detection of trophozoites or cysts in brain tissue. Corneal infections can be diagnosed by microscopic detection of amebae from corneal scrapings or histologic examination of corneal tissue (John 2001).
Acanthamoeba can be difficult to culture from infected animals (John 2001). Species of Acanthamoeba are diagnosed by analysis of isoenzyme patterns with electrophoresis and isoelectric focusing (Daggett et al. 1985), restriction enzyme analysis of mitochondrial DNA (Byers et al. 1983, Costas et al. 1983), as well as restriction endonuclease digestion of whole-cell DNA (McLaughlin et al. 1988).
In general, there is not a good correlation between pathogenicity and species of Acan- thamoeba (John 1993). However, there are genetic markers to help distinguish pathogenic from nonpathogenic strains of Acanthamoeba (Howe et al. 1997).
population effects Although numerous wild and domestic animals are susceptible to infection, there are no reported population effects of any significance (John 2001).
special problems Acanthamoeba spp. can serve as host for a number of pathogenic agents, and enhance their survival in soil or water. The capacity for some bacteria not normally considered soil and water organisms to survive in the environment for extended periods is influenced by their ability to successfully infect free-living Acanthamoeba. After being ingested by amebae, the bacteria grow, multiply, and eventually lyse the amebae and are released back into the environment. In addition to serving as a host for the soil saprophyte Listeria monocytogenes (Ly and Muller 1990), this unique relationship has been linked with extended environmental survival for Francisella tularensis (Abd et al. 2003), Salmonella enterica (Tezcan-Merdol et al. 2004), Chlamydia pneumoniae (Essig et al. 1997), Pasteurella multo- cida (Hundt and Ruffolo 2005), Mycobacterium avium (Cirillo et al. 1997, Steinert et al. 1998), and Mycobacterium leprae (Jadin 1975), among others. Thus, their extended capacity to survive in the environment by parasitizing amebae complicates the development of strategies to control these bacteria. There also is evidence that Acan- thamoeba can serve as suitable hosts for echoviruses and polioviruses (Danes and Cerva 1981).
control All work with prevention and control has been directed toward human cases.
There is no known satisfactory treatment for primary or granulomatous amebic meningoencephalitis (John 2001). There are several treatments for corneal infections, including topical treatments, epithelial debridement, and systemic ketoconazole (John 2001), as well as protective treatment of contact lenses (Hughes et al. 2003). No work has been reported with wildlife or domestic animals.
LITERATURE CITED
Abd, H., T. Johansson, I. Golovliov, G. Sandstrom, and M. Forsman. 2003. Survival and growth of Francisella tularensis in Acanthamoeba castel- lanii. Applied and Environmental Microbiology 69:600-606.
Acha, P. N., and B. Szyfres. 2003. Zoonoses and communicable diseases common to man and animals. Vol. III: Parasitoses. 3rd ed. Pan American Health Organization. Report No. 580.
Adl, S. M., A. G. B. Simpson, M. A. Farmer,
R. A. Andersen, O. R. Andersen, J. R. Barta,
S. S. Bowser, G. Brugerolle, R. A. Fensome,
S. Fredericq, T. Y. James, S. Karpov, P. Kugrens,
J. Krug, C. E. Lane, L. A. Lewis, J. Lodge,
D. H. Lynn, D. G. Mann, R. M. McCourt,
L. Mendoza, 0. Moestrup, S. E. Mozley- Standridge, T. A. Nerad, C. A. Shearer,
A. V. Smirnov, F. W. Spiegel, and M. F. J. R. Taylor. 2005. The new higher level classification of eukaryotes with emphasis on the taxonomy of protists. Journal of Eukaryotic Microbiology 52:399-451.
Allan, S. A. 2001. Biting flies (Class Insecta: Order Diptera). Pp. 18-45 in W. M. Samuel, M. J. Pybus, and A. A. Kocan (editors), Parasitic diseases of wild mammals. Iowa State University Press, Ames, IA.
Allsopp, R. 1984. Control of tsetse flies (Diptera: Glossinidae) using insecticides: A review and future prospects. Bulletin of Entomological Research 74:1-23.
Arkush, K. D., M. A. Miller, C. M. Leutenegger,
I. A. Gardner, A. E. Packham, A. R. Heckeroth, A. M. Tenter, B. C. Barr, and P. A. Conrad. 2003. Molecular and bioassay-based detection of Toxoplasma gondii oocyst uptake by mussels (Mytilus galloprovincialis). International Journal for Parasitology 33:1087-1097.
Ashcroft, M. T. 1959. The importance of African wild animals as reservoirs of trypanosomes. East African Medical Journal 36:289-297.
Atkinson, C. T. 1999. Hemosporidiosis. Pp. 193-199 in M. Friend and J. C. Franson (editors), Field manual of wildlife diseases: General field procedures and diseases of birds. U.S. Geological Survey, Biological Resources Division, Washington, DC.
Atkinson, C. T., R. J. Dusek, and J. K. Lease. 2001a. Serological responses and immunity to superinfection with avian malaria in experimentally- infected Hawaii amakihi. Journal of Wildlife Diseases 37:20-27.
Atkinson, C. T., R. J. Dusek, K. L. Woods, and W. M. Iko. 2000. Pathogenicity of avian malaria in experimentally-infected Hawaii amakihi. Journal of Wildlife Diseases 36:197-204.
Atkinson, C. T., D. J. Forrester, and E. C. Greiner. 1988. Pathogenicity of Haemoproteus meleagridis (Protozoa: Haemoproteidae) in experimentally infected domestic turkeys. Journal of Parasitology 74:228-239.
Atkinson, C. T., J. K. Lease, B. M. Drake, and N. P. Shema. 2001b. Pathogenicity, serological responses, and diagnosis of experimental and natural malarial infections in native Hawaiian thrushes. The Condor 103:209-218.
Atkinson, C. T., and C. van Riper III. 1991. Pathogenicity and epizootiology of avian haematozoa: Plasmodium, Leucocytozoon, and Haemoproteus. Pp. 19-48 in J. E. Loye and M. Zuk (editors), Bird-parasite interactions. Oxford University Press, Oxford, UK.
Atkinson, C. T., K. L. Woods, R. J. Dusek,
L. S. Sileo, and W. M. Iko. 1995. Wildlife disease and conservation in Hawaii: Pathogenicity of avian malaria (Plasmodium relictum) in experimentally infected iiwi (Vestiaria coccinea). Parasitology 111(Suppl.yS59-S69.
Baer, J. G. 1951. Ecology of animal parasites. University of Illinois Press, Urbana, IL.
Baer, J. G. 1971. Animal parasites. World University Library, McGraw-Hill Book Company, New York.
Baker, J. R. 1969. Trypanosomes of wild mammals in the neighbourhood of the Serengeti National Park. Pp. 147-155 in A. McDiarmid (editor), Diseases in free-living wild animals. Symposia of the Zoological Society of London No. 24. Academic Press, London, UK.
Baker, J. R. 1976. Biology of trypanosomes of birds. Pp. 131-174 in W. H. R. Lumsden and D. A. Evans (editors), Biology of the Kinetoplastida 1. Academic Press, London, UK.
Baker, J. R., R. Sachs, and I. Laufer. 1967. Trypanosomes of wild mammals in an area northwest of Serengeti National Park, Tanzania. Zeitschrift fur Tropenmedizin und Parasitologie 18:280-284.
Bardsley, J. E., and R. Harmsen. 1973. The trypanosomes of Anura. Advances in Parasitology 11:1-63.
Barnett, S. F., and D. W. Brocklesby. 1969. Some piroplasms of wild mammals. Pp. 159-176 in A. McDiarmid (editor), Diseases in free-living wild mammals. Symposia of the Zoological Society of London No. 24. Academic Press, London, UK.
Barta, J. R., D. S. Martin, P. A. Liberator,
M. Dashkevicz, J. W. Anderson, S. D. Feighner,
A. Elbrecht, A. Perkins-Barrow, M. C. Jenkins,
H. H. Danforth, M. D. Ruff, and
H. Profous-Juchelka. 1997. Phylogenetic relationships among eight Eimeria species infecting domestic fowl inferred using complete small subunit ribosomal DNA sequences. Journal of Parasitology 83:262-271.
Belova, L. M., and M. V. Krylov. 2003. The problems of evolution and phylogeny of coccidia with the eimerioran-type and isosporan-type of oocysts. Parazitologiya 37:324-332.
Bennett, G. F., M. A. Bishop, and M. A. Peirce. 1993. Checklist of the avian species of Plasmodium Marchiafava & Celli, 1885 (Apicomplexa) and their distribution by avian family and Wallacean life zones. Systematic Parasitology 26:171-179.
Bennett, G. F., J. R. Caines, and M. A. Bishop. 1988. Influence of blood parasites on the body mass of passeriform birds. Journal of Wildlife Diseases 24:339-343.
Bennett, G. F., M. A. Peirce, and R. A. Earle. 1994. An annotated checklist of the valid avian species of Haemoproteus, Leucocytozoon (Apicomplexa: Haemosporida) and Hepatozoon (Apicomplexa: Haemogregarinidae). Systematic Parasitology 29:61-73.
Bennett, G. F., M. Whiteway, and C. Woodworth- Lynas. 1982. A host-parasite catalogue of the avian haematozoa. Memorial University of Newfoundland Occasional Papers in Biology, No. 5, St. John's, Newfoundland, Canada.
Bert, E. 1619. An approved treatise of hawkes and hawking. Richard Moore Quaritch, London, England.
Bertram, B. C. R. 1973. Sleeping sickness survey in the Serengeti area (Tanzania) 1971. III: Discussion of the relevance of the trypanosome survey to the biology of large mammals in the Serengeti. Acta Tropica 30:36-48.
Boal, C. W., and R. W. Mannan. 1999. Comparative breeding ecology of Cooper's hawks in urban and exurban areas of southeastern Arizona. Journal of Wildlife Management 63:77-84.
Boal, C. W., R. W. Mannan, and K. S. Hudelson. 1998. Trichomoniasis in Cooper's hawks from Arizona. Journal of Wildlife Diseases 34:590-593.
BonDurant, R. H., and B. M. Honigberg. 1994. Trichomonads of veterinary importance.
Pp. 111-188 in J. P. Kreier (editor). Parasitic protozoa, Vol. 9. Academic Press, San Diego, CA.
BonDurant, R. H., and P. S. Wakenell. 1994. His- tomonas meleagridis and relatives. Pp. 189-206 in J. P. Kreier (editor), Parasitic protozoa, Vol. 9. Academic Press, San Diego, CA.
Boothroyd, J. C. 1985. Antigenic variation in African trypanosomes. Annual Review of Microbiology 39:475-502.
Borst, P. 1986. Discontinuous transcription and antigenic variation in trypanosomes. Annual Review of Biochemistry 55:701-732.
Bruckner, D. A., and J. A. Labarca. 1999. Leishmania and Trypanosoma. Pp. 1365-1373 in P. R. Murray, E. J. Baron, M. A. Pfaller, F. C. Tenover, and R. H. Yolken (editors), Manual of clinical microbiology. ASM Press, Washington, DC.
Bunbury, N., D. Bell, C. Jones, A. Greenwood, and P. Hunter. 2005. Comparison of the InPouch TF culture system and wet-mount microscopy for diagnosis of Trichomonas gallinae infections in the pink pigeon Columba mayeri. Journal of Clinical Microbiology 43:1005-1006.
Burrows, R. G., and M. A. Swerdlow. 1956. Entero- bius vermicularis as a probable vector of Dientamoeba fragilis. American Journal of Tropical Medicine and Hygiene 5:258-265.
Bush, A. O., J. C. Fernandez, G. W. Esch, and
J. R. Seed. 2001. Parasitism: The diversity and ecology of animal parasites. Cambridge University Press, Cambridge, UK.
Byers, T. J., S. A. Bogler, and L. L. Burianek. 1983. Analysis of mitochondrial DNA variation as an approach to systematic relationships in the genus Acanthamoeba. Journal of Protozoology 30:198-203.
Cabezon, O., A. R. Resendes, M. Domingo,
J. A. Raga, C. Agusti, F. Alegre, J. L. Mons,
J. P. Dubey, and S. Almeria. 2004. Seroprevalence of Toxoplasma gondii antibodies in wild dolphins from the Spanish Mediterranean coast. Journal of Parasitology 90:643-644.
Cannell, B. L., K. V. Krasnec, K. Campbell, H. I. Jones, R. D. Miller, and N. Stephens. 2013. The pathology and pathogenicity of a novel Haemoproteus spp. infection in wild little penguins (Eudyptula minor). Veterinary Parasitology. doi: 10.1016/j. vetpar.2013.1004.1025.
Carmichael, I. H., and E. Hobday. 1975. Blood parasites of some wild Bovidae in Botswana. Onderstepoort Journal of Veterinary Research 42:55-62.
Carmichael, J. 1933. The virus of rinderpest and its relation to Glossina morsitans (Weston). Bulletin of Entomological Research 24:337-342.
Cawthorn, R. J. 1993. Cyst-forming coccidia of raptors: Significant pathogens or not? Pp. 14-20 in P. T. Redig, J. E. Cooper, J. D. Remple, D. B. Hunter, and T. Hahn (editors), Raptor biomedicine. University of Minnesota Press, Minneapolis, MN.
Cedillos, R. A. 1975. Chagas' disease in El Salvador. Bulletin of the Pan American Health Organization 9:135-141.
Chandler, A. S., and C. P. Read. 1961. Introduction to parasitology. 10th ed. John Wiley and Sons, New York.
Cirillo, J. D., S. Falkow, L. S. Tompkins, and
L. E. Bermudez. 1997. Interaction of Mycobacterium avium with environmental amoebae enhances virulence. Infection and Immunity 65:3759-3767.
Clarke, J. E. 1964. Game elimination as a means of tsetse control with special reference to host preferences. The Puku 2:67-75.
Cockbill, G. F. 1967. The history and significance of trypanosomiasis problems in Rhodesia. Proceedings and Transactions of the Rhodesia Scientific Association 52:7-15.
Cole, R. A. 1999a. Renal coccidiosis. Pp. 215-218 in
M. Friend and J. C. Franson (editors), Field manual of wildlife diseases: General field procedures and diseases of birds. Biological Resources Division Information and Technology Report 1999-001. U.S. Geological Survey, Washington, DC.
Cole, R. A. 1999b. Trichomoniasis. Pp. 201-206 in M. Friend and J. C. Franson (editors), Biological Resources Division Information and Technology Report 1999-001. U.S. Geological Survey, Washington, DC.
Cole, R. A., and M. Friend. 1999. Miscellaneous parasitic diseases. Pp. 249-258 in M. Friend and J. C. Franson (editors), Field manual of wildlife diseases: General field procedures and diseases of birds. Biological Resources Division Information and Technology Report 1999-001. U.S. Geological Survey Washington, DC.
Cole, R. A., D. S. Lindsay, D. K. Howe, C. L. Roderick, J. P. Dubey, N. J. Thomas, and L. A. Baeten. 2000. Biological and molecular characterizations of Toxoplasma gondii strains obtained from southern sea otters (Enhydra lutris nereis). Journal of Parasitology 86:526-530.
Collins, E., and M. Aikawa. 1993. Plasmodia of nonhuman primates. Pp. 105-134 in J. P. Kreier (editor), Parasitic protozoa, Vol. 5. Academic Press, San Diego, CA.
Comes, A. M., J. F. Humbert, J. Cabaret, and L. Elard. 1996. Using molecular tools for diagnosis in veterinary parasitology. Veterinary Research 27:333-342.
Conrad, P. A., M. A. Miller, C. Kreuder, E. R. James, J. Mazet, H. Dabritz, D. A. Jessup, F. Gulland, and M. E. Grigg. 2005. Transmission of Toxoplasma: Clues from the study of sea otters as sentinels of Toxoplasma gondii flow into the marine environment. International Journal for Parasitology 35:1155-!!68.
Conti, J. A. 1993. Diseases, parasites and contaminants. Pp. 205-224 inT. S. Baskett, M. W. Sayre,
R. E. Tomlinson, and R. E. Mirarchi (editors), Ecology and management of the mourning dove. Stackpole Books, Harrisburg, PA.
Cook, R. S. 1971a. Haemoproteus. Pp. 300-308 in
J. W. Davis, R. C. Anderson, L. Karstad, and
D. O. Trainer (editors), Infectious and parasitic diseases of wild birds. Iowa State University Press, Ames, IA.
Cook, R. S. 1971b. Leucocytozoon Danilewsky 1890. Pp. 291-299 in J. W. Davis, R. C. Anderson,
L. Karstad, and D. O. Trainer (editors), Infectious and parasitic diseases of wild birds. Iowa State University Press, Ames, IA.
Cooper, J. E. 2002. Birds of prey: Health and disease. 3rd ed. Blackwell Science, Oxford, UK.
Cooper, J. E., and S. J. Petty. 1988. Trichomoniasis in free-living goshawks (Accipiter gentilis gentilis) from Great Britain. Journal of Wildlife Diseases 24:80-87.
Costas, M., and A. J. Griffiths. 1986. Physiological characterizations of Acanthamoeba strains. Journal of Protozoology 32:604-607.
Costas, M., S. W. Edwards, D. Lloyd, A. J. Griffiths, and G. Turner. 1983. Restriction enzyme analysis of mitochondrial DNA of members of the genus Acanthamoeba as an aid in taxonomy. FEMS (Federation of European Microbiology Societies) Microbiology Letters 17:231-234.
Cover, A. J., W. M. Harmon, and M. W. Thomas. 1994. A new method for the diagnosis of Trichomonas gallinae infection by culture. Journal of Wildlife Diseases 30:457-457.
Cox, F. E. G. 1991. Systematics of parasitic protozoa.
Pp. 55-80 in J. P. Kreier and J. R. Baker (editors), Parasitic protozoa. Academic Press, San Diego, CA.
Cox, F. E. G. 1993. Plasmodia of rodents. Pp. 49-105 in J. P. Kreier (editor), Parasitic protozoa, Vol. 5. Academic Press, San Diego, CA.
Craig, T. M. 2001. Hepatozoon spp. and hepatozoonosis. Pp. 462-468 in W. M. Samuel,
M. J. Pybus, and A. A. Kocan (editors), Parasitic diseases of wild mammals. Iowa State University Press, Ames, IA.
Critcher, S. 1950. Renal coccidiosis in Pea Island Canada geese. Wildlife of North Carolina 14:14-15.
Cruikshank, J. J., D. M. Haines, N. C. Palmer, and
D. J. S. Aubin. 1990. Cysts of a Toxoplasma-like organism in an Atlantic bottlenose dolphin. Canadian Veterinary Journal 31:213-215.
Daggett, P.-M., D. Lipscomb, T. K. Sawyer, and
T. A. Nerad. 1985. A molecular approach to the phylogeny of Acanthamoeba. BioSystems ⅛399-405.
Danes, L., and L. Cerva. 1981. Survival of polioviruses and echoviruses in Acanthamoeba castellanii cultivated in vitro. Journal of Hygiene, Epidemiology, Microbiology and Immunology 25:169-174.
Donelson, J. E., and M. J. Turner. 1985. How the trypanosome changes its coat. Scientific American 252:44-51.
Dubey, J. P. 1993. Toxoplasma, Neospora, Sarcocystis, and other tissue cyst-forming coccidia of humans and animals. Pp. 1-158 in J. P. Kreier (editor), Parasitic protozoa, Vol. 6. Academic Press, San Diego, CA.
Dubey, J. P. 2001. Oocyst shedding by cats fed isolated bradyzoites and comparison of infectivity of bradyzoites of the VEG strain Toxoplasma gondii to cats and mice. Journal of Parasitology 87:215-219.
Dubey, J. P. 2004. Toxoplasmosis: A waterborne zoonosis. Veterinary Parasitology 126:57-72.
Dubey, J. P., and C. P. Beattie. 1988. Toxoplasmosis of animals and man. CRC Press, Boca Raton, FL.
Dubey, J. P., and J. L. Carpenter. 1993a. Histologically confirmed clinical toxoplasmosis in cats: 100 cases (1952-1990). Journal of the American Veterinary Medical Association 203:1556-1566.
Dubey, J. P., and J. L. Carpenter. 1993b. Neonatal toxoplasmosis in littermate cats. Journal of the American Veterinary Medical Association 203:1546-1549.
Dubey, J. P., P. A. Fair, G. D. Bossart, D. Hill, R. Fayer, C. Sreekumar, O. C. H. Kwok, and P. Thulliez. 2005. A comparison of several serologic tests to detect antibodies of Toxoplasma gondii in naturally exposed bottlenose dolphins (Tursiops truncatus). Journal of Parasitology 91:1074-1081.
Dubey, J. P., and J. K. Frenkel. 1972. Cyst-induced toxoplasmosis in cats. Journal of Protozoology 19:155-177.
Dubey, J. P., R. P. Lipscomb, and M. Mense. 2004. Toxoplasmosis in an elephant seal (Mirounga angustirostris). Journal of Parasitology 90:410-411.
Dubey, J. P., N. L. Miller, and D. K. Frenkel. 1970. Toxoplasma gondii life cycle in cats. Journal of the American Veterinary Medical Association 157:1767-1770.
Dubey, J. P., and K. Odening. 2001. Toxoplasmosis and related infections. Pp. 478-519 in W. M. Samuel, M. J. Pybus, and A. A. Kocan (editors), Parasitic diseases of wild mammals. Iowa State University Press, Ames, IA.
Dubey, J. P., R. Zarnke, N. J. Thomas, S. K. Wong, W. V. Bonn, M. Briggs, J. W. Davis, R. Ewing, M. Mense, O. C. H. Kwok, S. Romand, and P. Thulliez. 2003. Toxoplasma gondii, Neospora caninum, Sarcocystis neurona, and Sarcocystis canis-like infections in marine mammals. Veterinary Parasitology 116:275-296.
Duke, H. L. 1919. An enquiry into the relations of Glossina morsitans and ungulate game, with special reference to rinderpest. Bulletin of Entomological Research 10:7-20.
Dunsmore, J. D. 1971. A study of the biology of the wild rabbit in climatically different regions in eastern Australia. IV: The rabbit in the south coastal region of New South Wales, an area in which parasites appear to exert a populationregulating effect. Australian Journal of Zoology 19:355-370.
Duszynski, D. W., and S. J. Upton. 2001. Cyclospora, Eimeria, Isospora, and Cryptosporidium spp. Pp. 416-459 in W. M. Samuel, M. J. Pybus, and A. A. Kocan (editors), Parasitic diseases of wild mammals. Iowa State University Press, Ames, IA.
Du Toit, R. 1954. Trypanosomiasis in Zululand and the control of tsetse flies by chemical means. Onderstepoort Journal of Veterinary Research 26:317-387.
Eichinger, D. 2001. A role for a galactose lectin and its ligands during encystment of Entamoeba. Journal of Eukaryotic Microbiology 48:17-21.
Essig, A., M. Heinemann, U. Simnacher, and R. Marre. 1997. Infection of Acanthamoeba castellanii by Chlamydia pneumoniae. Applied and Environmental Microbiology 63:1396-1399.
Estes, J. A., B. B. Hatfield, K. Ralls, and J. Ames. 2003. Causes of mortality in California sea otters during periods of population growth and decline. Marine Mammal Science 19:198-216.
Farr, M. M. 1954. Renal coccidiosis of Canada geese. Journal of Parasitology 40(Suppl.)46.
Fayer, R., J. P. Dubey, and D. S. Lindsay. 2004. Zoonotic protozoa: From land to sea. Trends in Parasitology 20:531-536.
Ford, J. 1971. The role of trypanosomiasis in African Ecology. Clarendon Press, Oxford, UK.
Frank, W. 1976. Parasitologie. Verlag Eugen Ulmer, Stuttgart, Germany.
Franker, C. K., and J. P. Doll. 1964. Experimental histomoniasis in gnotobiotic turkeys. II: Effects of some cecal bacteria on pathogenesis. Journal of Parasitology 50:636-640.
Fraser, C. M., and A. Mays. 1986. The Merck veterinary manual. 6th ed. Merck & Co., Inc., Rahway, NJ.
Frenkel, J. K., A. Ruiz, and M. Chinchilla. 1975. Soil survival of Toxoplasma oocysts in Kansas and Costa Rica. American Journal of Tropical Medicine and Hygiene 24:439-443.
Friend, M., and J. C. Franson. 1999. Intestinal coccidiosis. Pp. 207-213 in M. Friend and J. C. Franson (editors), Field manual of wildlife diseases: General field procedures and diseases of birds. Biological Resources Division,
U.S. Geological Survey, Washington, DC.
Gajadhar, A. A., L. Measures, L. B. Forbes, C. Kapel, and J. P. Dubey. 2004. Experimental Toxoplasma gondii infection in grey seals (Halichoerus grypus). Journal of Parasitology 90:255-259.
Garcia, L. S., and D. A. Bruckner. 1997. Diagnostic medical parasitology. 3rd ed. American Society for Microbiology, Washington, DC.
Garnham, P. C. C. 1966. Malaria parasites and other Haemosporidia. Blackwell Scientific, Oxford, UK.
Geigy, R., and M. Kauffman. 1973. Sleeping sickness survey in the Serengeti area (Tanzania) 1971. I: Examination of large mammals for trypanosomiasis. Acta Tropica 30:12-23.
Geigy, R., M. Kauffman, and R. Beglinger. 1967.
A survey of wild animals as potential reservoirs of trypanosomiasis in the Ulanga District (Tanzania). Acta Tropica 24:97-108.
Gerber, L. R., M. T. Tinker, D. F. Doak, and
J. A. Estes. 2004. Mortality sensitivity in life-stage simulation analysis: A case study of southern sea otters. Ecological Applications 14:1554-1565.
Glover, P. E. 1965. A review of recent knowledge on the relationship between the tsetse fly and its vertebrates hosts. Fauna Preservation Society, London, UK.
Graham, P. 1964. Destruction of birds and other wildlife by dieldrex spraying against tsetse fly in Bechuanaland. Arnoldia (Rhodesia) 1:1-4.
Graybill, H. W., and T. Smith. 1920. Production of fatal blackhead in turkeys by feeding embryo- nated eggs of Heterakis papillosa. Journal of Experimental Medicine 31:647-655.
Grigg, M. E., S. Bonnefoy, A. B. Hehl, Y. Suzuki, and J. C. Boothroyd. 2001. Success and virulence in Toxoplasma as the result of sexual recombination between two distinct ancestries. Science 294:161-165.
Gyening, K. O. 1990. Organizational perceptions and current approaches to trypanosomiasis problems undertaken by FAO: Consultation on trypanosomiasis research and programmes in Africa, 30 April-4 May 1990. International Laboratory for Research on Animals Diseases, Nairobi, Kenya.
Gylstorff, I., and F. Grimm. 1987. Vogelkrankheiten. Verlag Eugen Ulmer, Stuttgart, Germany.
Hanson, H. C., N. D. Levine, and V. Ivens. 1957.
Coccidia (Protozoa: Eimeriidae) of North American wild geese and swans. Canadian Journal of z°°l°gy 35:715-733.
Harmon, W. M., W. A. Clark, A. C. Hawbecker, and M. Stafford. 1987. Trichomonas gallinae in colum- biform birds from the Galapagos Islands. Journal of Wildlife Diseases 23:492-494.
Heisch, R. G. 1963. Presence of trypanosomes in bush babies after eating infected rats. Nature (London) 169:118.
Herman, C. M. 1969. Blood protozoa of free-living birds. In A. McDiarmid (editor), Diseases in free- living wild animals. Symposia of the Zoological Society of London No. 24. Academic Press, London, UK.
Hill, D. E., S. Chirukandoth, and J. P. Dubey. 2005. Biology and epidemiology of Toxoplasma gondii in man and animals. Animal Health Research Reviews 6:41-61.
Hoare, C. A. 1966. The classification of mammalian trypanosomes. Ergebnisse der Mikrobiologie, Immunitatsforschung und experimentellen Therapie 39:43-57.
Hoare, C. A., and F. G. Wallace. 1966. Developmental stages of trypanosomatid flagellates: A new terminology. Nature 212:1385-1386.
Hofle, U., C. Gortazar, J. A. Ortiz, B. Knispel, and
E. F. Kaleta. 2004. Outbreak of trichomoniasis in a woodpigeon (Columba palumbus) wintering roost. European Journal of Wildlife Research 50:73-77.
Honnold, S. P., R. Braun, D. P. Scott, C. Sreekumar, and J. P. Dubey. 2005. Toxoplasmosis in a Hawaiian monk seal (Monachus schauinslandi). Journal of Parasitology 91:695-697.
Howe, D. K., M. H. Vodkin, R. J. Novak,
G. S. Visvesvara, and G. L. McLaughlin. 1997. Identification of two genetic markers that distinguish pathogenic and nonpathogenic strains of Acanthamoeba spp. Parasitology Research 83:345-348.
Huff, C. G. 1965. Susceptibility of mosquitoes to avian malaria. Experimental Parasitology 16:107-132.
Hughes, R., P. W. Andrew, and S. Kilvington. 2003. Enhanced killing of Acanthamoeba cysts with a plant peroxidase-hydrogen peroxide-halide antimicrobial system. Applied and Environmental Microbiology 69:2563-2567.
Hundt, M. J., and C. G. Ruffolo. 2005. Interaction of Pasteurella multocida with free-living amoeba. Applied and Environmental Microbiology 7E5458-5464.
Inskeep, W., 2nd, C. H. Gardiner, R. K. Harris, J. P. Dubey, and R. T. Goldston. 1990. Toxoplasmosis in Atlantic bottle-nosed dolphins (Tursiops truncatus). Journal of Wildlife Diseases 26:377-382.
Jadin, J. B. 1975. Amibes “Limax” vecteurs possibles des Mycobacteries et de Mycobacterium leprae. Acta Leprologica 59-60:57-67.
Jakob, W., and H. H. Wesemeier. 1995. Intestinal inflammation associated with flagellates in snakes. Journal of Comparative Pathology 112:417-421.
Jardine, J. E., and J. P. Dubey. 2002. Congenital toxoplasmosis in a Indo-Pacific bottlenose dolphin (Tursiops aduncus). Journal of Parasitology 88:197-199.
Jawara, A. S. D. K. 1990. Animal disease as a factor limiting economic development in Africa. Cornell Veterinarian 80:17-25.
Jessup, D. A., M. Miller, J. Ames, M. Harris,
C. Kreuder, P. A. Conrad, and J. A. K. Mazet. 2004. Southern sea otter as a sentinel of marine ecosystem health. EcoHealth 1:239-245.
Jindrichova, J., K. Kramarova, B. Rosicky, J. Jira, and A. Simko. 1975. The cat as a possible source of Toxoplasma infection for man. Folia Parasito- logica 22:309-315.
John, D. T. 1993. Opportunistically pathogenic free- living amebae, Vol. 3. Pp. 143-246 in J. P. Kreier and J. R. Baker (editors), Parasitic protozoa. Academic Press, San Diego, CA.
John, D. T. 2001. Opportunistic amoebae.
Pp. 460-462 in W. M. Samuel, M. J. Pybus, and
A. A. Kocan (editors), Parasitic diseases of wild mammals. Iowa State University Press, Ames, IA.
Jordan, A. M. 1974. Recent developments in the ecology and methods of control of tsetse flies (Glossina spp.) (Dipt. Glossinidae): A review. Bulletin of Entomological Research 63:361-399.
Julian, R. J., and D. E. Galt. 1980. Mortality in muscovy ducks (Cairina moschata) caused by Haemoproteus infection. Journal of Wildlife Diseases 16:39-44.
Kakoma, 1., and H. Mehlhorn. 1994. Babesia of domestic animals. Pp. 141-216 in J. P. Kreier (editor), Parasitic protozoa, Vol. 7. Academic Press, San Diego, CA.
Keeton, W. T., and J. L. Gould. 1993. Biological science, Vol. 2. 5th ed. W. W. Norton & Company, New York.
Kellogg, F. E., T. H. Eleazer, and T. R. Colvin. 1972. Transmission of blackhead from jungle fowl to turkey. Proceedings of the Annual Conference of the Southeastern Association of Fish and Wildlife Agencies 32:378-379.
Kellogg, F. E., and W. M. Reid. 1970. Bobwhites as possible reservoir hosts for blackhead in wild turkeys. Journal of Wildlife Management 34U55→59.
Kemp, R. L. 1974. The failure of Histomonas melea- gridis to establish in germ-free ceca in normal poults. Avian Diseases 18:452-455.
Kemp, R. L., and J. C. Franson. 1975. Transmission of Histomonas meleagridis to domestic fowl by means of earthworms recovered from pheasant yard soil. Avian Diseases 19:741-744.
Kimura, M., A. A. Dhondt, and I. J. Lovette. 2006. Phylogeographic structuring of Plasmodium lineages across the North American range of the house finch (Carpodacus mexicanus). Journal of Parasitology 92:1043-1049.
Kocan, A. A. 2001a. Amebiasis, balantidiasis, and enteric trichomoniasis. Pp. 397-399 in W. M. Samuel, M. J. Pybus, and A. A. Kocan (editors), Parasitic diseases of wild mammals. Iowa State University Press, Ames, IA.
Kocan, A. A. 2001b. Blood-inhabiting protozoan parasites. Pp. 520-524 in W. M. Samuel, M. J. Pybus, and A. A. Kocan (editors), Parasitic diseases of wild mammals. Iowa State University Press, Ames, IA.
Kocan, A. A., and K. A. Waldrup. 2001. Piroplasms (Theileria spp., Cytauxzoon spp., and Babesia spp.). Pp. 524-536 in W. M. Samuel, M. J. Pybus, and A. A. Kocan (editors), Parasitic diseases of wild mammals. Iowa State University Press, Ames, IA.
Kreuder, C., M. A. Miller, D. A. Jessup,
L. J. Lowenstine, M. D. Harris, J. A. Ames,
T. E. Carpenter, P. A. Conrad, and J. A. K. Mazet.
2003. Patterns of mortality in southern sea otters (Enhydra lutris nereis) from 1998-2001. Journal of Wildlife Diseases 39:495-509.
Krone, O., R. Altenkamp, and N. Kenntner. 2005. Prevalence of Trichomonas gallinae in northern goshawks from the Berlin area of northeastern Germany. Journal of Wildlife Diseases 41:304-309.
Kudo, R. R. 1966. Protozoology. Charles C. Thomas, New York.
Kulda, J., and E. Nohynkova. 1994. Giardia in humans and animals. Pp. 225-422 in J. P. Kreier (editor), Parasitic protozoa, Vol. 10. Academic Press, San Diego, CA.
Lainson, R. 1982. Leishmania parasites of mammals in relation to human health. Pp. 137-179 in
M. A. Edwards and U. McDonnell (editors), Animal disease in relation to animal conservation. Symposia of the Zoological Society of London No. 50. Academic Press, London, UK.
Lambourn, D. M., S. J. Jeffries, and J. P. Dubey. 2001. Seroprevalence of Toxoplasma gondii in harbor seals (Phoca vitulina) in southern Puget Sound, Washington. Journal of Parasitology 87:1196-1197.
Leber, A. L., and S. M. Novak. 1999. Intestinal and urogenital amebae, flagellates, and ciliates. Pp. 1391-1405 in P. R. Murray, E. J. Baron, M. A. Pfaller, F. C. Tenover, and R. H. Yolken (editors), Manual of clinical microbiology. ASM Press, Washington, DC.
Lee, D. H. 1970. The structure and development of Histomonas meleagridis (Mastigamoebidae: Protozoa) in the female reproductive tract and its intermediate host, Heterakis gallinarum (Nematoda). Parasitology 59:877-884.
Lehmann, T., P. L. Marcet, D. H. Graham, E. R. Dahl, and J. P. Dubey. 2006. Globalization and the population structure of Toxoplasma gondii. Proceedings of the National Academy of Sciences of the United States of America 103:11423-11428.
Leighton, F. A., and A. A. Gajadhar. 2001.
Besnoitia spp. and besnoitiosis. Pp. 468-478 in W. M. Samuel, M. J. Pybus, and A. A. Kocan (editors), Parasitic diseases of wild mammals. Iowa State University Press, Ames, IA.
Lindsay, D. S., M. V. Collins, S. M. Mitchell,
R. A. Cole, G. J. Flick, C. N. Wetch, A. Lindquist, and J. P. Dubey. 2003. Sporulation and survival of Toxoplasma gondii oocysts in seawater. Journal of Eukaryotic Microbiology 50:687-688.
Lindsay, D. S., M. V. Collins, S. M. Mitchell,
C. N. Wetch, C. N. Wetch, A. C. Rosypal, G. J. Flick, A. M. Zajac, A. Lindquist, and J. P. Dubey. 2004. Survival of Toxoplasma gondii oocysts in eastern oysters (Crassostrea virginica). Journal of Parasitology 90:1054-1057.
Lindsay, D. S., K. K. Phelps, S. A. Smith, G. Flick,
S. S. Sumner, and J. P. Dubey. 2001a. Removal of Toxoplasma gondii oocysts from sea water by eastern oysters (Crassostrea virginica). Journal of Eukaryotic Microbiology 48:i97s-i98s.
Lindsay, D. S., N. J. Thomas, A. C. Rosypal, and J. P. Dubey. 2001b. Dual Sarcocystis neurona and Toxoplasma gondii infection in a northern sea otter from Washington State, USA. Veterinary Parasitology 97:319-327.
Lindsay, D. S., and K. S. Todd. 1993. Coccidia of mammals. Pp. 89-131 in J. P. Kreier (editor), Parasitic protozoa, Vol. 4. Academic Press, San Diego, CA.
Logan-Henfrey, L. L., P. R. Gardiner, and
M. M. Mahmoud. 1992. Animal trypanosomiasis in sub-Sahara Africa. Pp. 157-276 in J. P. Kreier and J. R. Baker (editors), Parasitic protozoa, Vol. 2. Academic Press, San Diego, CA.
Lom, J. 1979. Biology of the trypanosomes and trypanoplasms of fish. Pp. 269-337 in W. H. R. Lumsden and D. A. Evans (editors), Biology of the Kinetoplastida 2. Academic Press, London, UK.
Long, P. F. 1993. Avian coccidiosis. Pp. 1-88 in J. P. Kreier (editor), Parasitic protozoa, Vol. 4. Academic Press, San Diego, CA.
Lopez-Antunano, F. J., and G. A. Schmunis. 1993. Plasmodia of humans. Pp. 135-266 in J. P. Kreier (editor), Parasitic protozoa, Vol. 5. Academic Press, San Diego, CA.
Lucius, R., and B. Loos-Frank. 1997. Parasitologie. Spektrum Akademischer Verlag GmbH, Heidelberg, Germany.
Lund, E. E., and A. M. Chute. 1971. Bobwhite, Colinus virginianus, as host for Heterakis and Histomonas. Journal of Wildlife Diseases 7:70-75.
Lund, E. E., E. E. Wehr, and D. J. Ellis. 1966. Earthworm transmission of Heterakis and Histomonas to turkeys and chickens. Journal of Parasitology 52:899-902.
Ly, T. M., and H. E. Muller. 1990. Ingested Listeria monocytogenes survive and multiply in protozoa. Journal of Medical Microbiology 33:51-54.
Ma, P., G. S. Visvesvara, A. J. Martinez,
F. H. Theodore, P. M. Daggett, and T. K. Sawyer. 1990. Naegleria and Acanthamoeba infections: review. Review of Infectious Diseases 12:490-513.
Mackenstedt, U., M. Gauer, H. Mehlhorn,
E. Schein, and S. Hauschild. 1990. Sexual cycle of Babesia divergens confirmed by DNA measurements. Parasitology Research 76:199-206.
Martinez, A. J., and G. S. Visvesvara. 1997. Free- living, amphizoic and opportunistic amebas. Brain Pathology 7:583-598.
Martinez-Paloma, A. 1993. Parasitic amoebas in the intestinal tract. Pp. 65-141 in J. P. Kreier and J. R. Baker (editors), Parasitic protozoa. Academic Press, New York.
Massie, G., and M. Black. 2008. Unraveling the mystery of the kitty litter parasite in marine mammals. Science Daily, American Society for Microbiology (June 5)..
Matthiessen, P. 1985. Contamination of wildlife with DDT insecticide residues in relation to tsetse fly control operations in Zimbabwe. Environmental Pollution Series B: Chemical and Physical 10:189-211.
Matthiessen, P., and B. Douthwaite. 1985. The impact of tsetse fly control campaigns on African wildlife. Oryx 19:202-209.
McDougald, L. R. 1991. Other protozoal diseases of the intestinal tract. Pp. 804-809 in B. W. Calnek, H. J. Barnes, C. W. Beard, W. M. Reid, and H. W. Yoder (editors), Diseases of poultry. Iowa State University Press, Ames, IA.
McDougald, L. R., and W. M. Reid. 1978. Histomonas meleagridis and its relatives. Pp. 139-161 in J. P. Kreier (editor), Parasitic protozoa, Vol. II. Academic Press, New York.
McLaughlin, G. L., F. H. Brandt, and G. S. Visvesvara. 1988. Restriction fragment length polymorphisms of the DNA of selected Naegleria and Acanthamoeba amebae. Journal of Clinical Microbiology 26:1655-1658.
Measures, L. N., J. P. Dubey, P. Labelle, and
D. Martineau. 2004. Seroprevalence of Toxoplasma gondii in Canadian pinnipeds. Journal of Wildlife Diseases 40:294-300.
Meerovitch, E. 1958. Some biological requirements and host parasite relations of Entamoeba invadens. Canadian Journal of Zoology 36:513-523.
Mehlhorn, H., W. Raether, E. Schein, M. Weber, and M. Uphoff. 1986. Licht und elektronenmi- krskopische Untersuchungen zum Entwicklung- szyklus und Einfluss von Pentamichin auf die Morphologie der intraerythrozytaren Stadien von Babesia microti. Deutsche Tierartzliche Wochenschrift 93:377-464.
Mehlhorn, H., E. Schein, and J. S. Ahmed. 1994. Theileria. Pp. 217-304 in J. P. Kreier (editor), Parasitic protozoa, Vol. 7. Academic Press, San Diego, CA.
Miller, M. A., I. A. Gardner, C. Kreuder,
D. M. Paradies, K. R. Worcester, D. A. Jessup,
E. Dodd, M. D. Harris, J. A. Ames,
A. E. Packham, and P. A. Conrad. 2002a.
Coastal freshwater runoff is a risk factor for Toxoplasma gondii infection of southern sea otters (Enhydra lutris nereis). International Journal for Parasitology 32:997-1006.
Miller, M. A., I. A. Gardner, A. Packham,
J. K. Mazet, K. D. Hanni, D. Jessup, J. Estes,
R. Jameson, E. Dodd, B. C. Barr, L. J. Lowenstine,
F. M. Gulland, and P. A. Conrad. 2002b. Evaluation of an indirect fluorescent antibody test (IFAT) for demonstration of antibodies to Toxoplasma gondii in the sea otter (Enhydra lutris). Journal of Parasitology 88:594-599.
Miller, M. A., M. E. Grigg, C. Kreuder, E. R. James, A. C. Melli, P. R. Crosbie, D. A. Jessup, J. C. Boothroyd, D. Brownstein, and P. A. Conrad.
2004. An unusual genotype of Toxoplasma gondii is common in California sea otters (Enhydra lutris nereis) and is a cause of mortality. International Journal for Parasitology 34:275-284.
Mitchell, S., and A. Sinai. 2006. Toxoplasma gondii: Presentations at the IX International Workshop on Opportunistic Protists and the International Society of Protistologists 57th Annual Meetings. Journal of Eukaryotic Microbiology 53^159.
Moloo, S. K., G. J. Losos, and S. S. Katuza. 1973. Transmission of Trypanosoma brucei to cats and dogs by feeding on infected goats. Annals of Tropical Medicine and Parasitology 67:331-334.
Moltmann, U. G., H. Mehlhorn, E. Schein,
G. Rehbein, W. P. Voigt, and E. Zweygarth. 1983. Fine structure of Babesia equi (Laveran 1901) within lymphocytes and erythrocytes of horses: An in vivo and in vitro study. Journal of Parasitology 69:111-120.
Molyneux, D. H. 1982. Trypanosomes, trypanosomiasis and tsetse control: Impact on wildlife and its conservation. Pp. 29-55 in M. A. Edwards and U. McDonnell (editors), Animal disease in relation to animal conservation. Symposia of the Zoological Society of London No. 50. Academic Press, London, UK.
Ockert, G., and T. Schmidt. 1976. On the epidemiology of Dientamoeba fragilis Jepps and Dobell 1918. 4th communication: Evidence of Dientamoeba fragilis in Enterobius eggs using isoelectric point determination [in German]. Journal of Hygiene Epidemiology Microbiology and Immunology (Prague) 20:76-81.
Olsen, O. W. 1974. Animal parasites: Their life cycles and ecology. 3rd ed. University Park Press, Baltimore, MD.
Olson, M. E., and A. G. Buret. 2001. Giardia and giardiasis. Pp. 399-416 in W. M. Samuel, M. J. Pybus, and A. A. Kocan (editors), Parasitic diseases of wild mammals. Iowa State University Press, Ames, IA.
Ostrand, W. D., J. A. Bissonette, and M. R. Conover. 1995. Trichomoniasis as a factor in mourning dove population decline in Fillmore, Utah. Journal of Wildlife Diseases 31:87-89.
Patterson, D. J. 1999. The diversity of eukaryotes. American Naturalist 65^96-S124.
Patterson, D. J. 2000. Changing views on protistan systematics: The taxonomy of protozoa—an overview. Pp. 2-9 in J. J. Lee, G. F. Leedale, and P. Bradbury (editors), An illustrated guide to the protozoa: Organisms traditionally referred to as protozoa, or newly discovered groups. Society of Protozoologists, Lawrence, KS.
Pays, E., and M. Steinert. 1988. Control of antigen gene expression in African trypanosomes. Annual Review of Genetics 22:107-126.
Peirce, M. A. 2000. A taxonomic review of avian piroplasms of the genus Babesia Starcovici, 1893 (Apicomplexa: Piroplasmorida: Babesidae). Journal of Natural History 34:317-332.
Peirce, M. A. 2005. Pathogenic subspecies of Plasmodium relictum found in African birds. Veterinary Record 156:328.
Pepler, D., and E. E. Oettle. 1992. Trichomonas gallinae in wild raptors on the Cape Peninsula. South African Journal of Wildlife Research 22:87-88.
Philippa, J. D. W., F. A. Leighton, P. Y. Daoust,
O. Nielsen, H. Schwantje, T. Shury,
R. v. Herwijnen, B. E. E. Martina, T. Kuiken, M. W. G. V. d. Bildt, and A. D. M. E. Osterhaus. 2004. Antibodies to selected pathogens in free-ranging terrestrial carnivores and marine mammals in Canada. Veterinary Record I55:i35-i40.
Pokras, M. A., E. B. Wheeldon, and C. J. Sedgwick.
i993. Trichomoniasis in owls: Report on a number of clinical cases and a survey of the literature. Pp. 88-91 in P. T. Redig, J. E. Cooper, J. D. Remple, D. B. Hunter, and T. Hahn (editors), Raptor biomedicine. University of Minnesota Press, Minneapolis, MN.
Quortrup, E. R., and J. E. Shillinger. 1941. 3,000 wild bird autopsies on western lake areas. Journal of the American Veterinary Medical Association 99:382-387.
Ratcliffe, H. L., and Q. M. Geiman. 1938.
Spontaneous and experimental amebic infections in reptiles. Archives of Pathology 25:160-184.
Renold, C., A. Sugar, J. P. Chave, L. Perrin,
J. Delavelle, G. Pizzolato, P. Burkhardt,
V. Gabriel, and B. Hirschel. 1992. Toxoplasma encephalitis in patients with the acquired immunodeficiency syndrome. Medicine 71.
Resendes, A. R., S. Almeria, J. P. Dubey, E. Obon,
C. Juan-Salles, E. Degollada, F. Alegre,
O. Cabezon, S. Pont, and M. Domingo. 2002. Disseminated toxoplasmosis in a Mediterranean pregnant Risso's dolphin (Grampus griseus) with transplacental fetal infection. Journal of Parasitology 88:1029-1032.
Roberts, L. S., and J. Janovy. 2000. Gerald D.
Schmidt and Larry S. Roberts' Foundations of parasitology. 6th ed. McGraw-Hill, Boston, MA.
Rosenfield, R. N., J. Bielefeldt, L. J. Rosenfield,
S. J. Taft, R. K. Murphy, and A. C. Stewart. 2002. Prevalence of Trichomonas gallinae in nestling Cooper's hawks among three North American populations. Wilson Bulletin 114:145-147.
Ruiz, A., and J. K. Frenkel. 1980. Toxoplasma gondii in Costa Rican cats. American Journal of Tropical Medicine and Hygiene 29:1150-1160.
Sanger, V. L. 1971. Toxoplasmosis. Pp. 313-316 in J. W. Davis, R. C. Anderson, L. Karstad, and
D. O. Trainer (editors), Infectious and parasitic diseases of wild birds. Iowa State University Press, Ames, IA.
Sawyer, T. K., G. S. Visvesvara, and B. A. Harke. 1976. Pathogenic amebas from brackish and ocean sediments with a description of Acanthamoeba hatchetti, n. sp. Science i96:i324-i325.
Slapeta, J. R., D. Modry, J. Votypka, M. Jirku, J. Lukes, and B. Koudela. 2003. Evolutionary relationships among cyst-forming coccidia Sarco- cystis spp. (Alveolata: Apicomplexa: Coccidea) in endemic African tree vipers and perspective for evolution of heteroxenous life cycle. Molecular Phylogenetics and Evolution 27:464-475.
Sleigh, M. A. 1991. The nature of protozoa. Pp. 1-53 in J. P. Kreier and J. R. Baker (editors), Parasitic protozoa. Academic Press, San Diego, CA.
Sousa, O. E., and C. M. Johnson. 1973. Prevalence of Trypanosoma cruzi and Trypanosoma rangeli in triatomines (Hemiptera: Reduviidae) collected in the Republic of Panama. American Journal of Tropical Medicine and Hygiene 22:18-23.
Springer, W. T., J. Johnson, and W. M. Reid. 1969. Transmission of histomoniasis with male Heterakisgallinarum (Nematoda). Parasitology 59:40i-405.
Springer, W. T., J. Johnson, and W. M. Reid. i970. Histomoniasis in gnotobiotic chickens and turkeys: Biological aspects of the role of bacteria in the etiology. Experimental Parasitology 28:383-392.
Stabler, R. M. 1954. Trichomonas gallinae: A review. Experimental Parasitology 3:368-402.
Stabler, R. M. 1975. Personal letter, 21 May 1975. In R. G. Botzler (editor). Humboldt State University, Arcata, CA.
Steenbergen, J. N., J. D. Nosanchuk, S. D. Malliaris, and A. Casadevall. 2004. Interaction of Blastomyces dermatitidis, Sporothrix schenckii, and Histoplasma capsulatum with Acanthamoeba castellanii. Infection and Immunity 72:3478-3488.
Steinert, M., K. Birkness, E. White, B. Fields, and
F. Quinn. i998. Mycobacterium avium bacilli grow saprozoically in coculture with Acanthamoeba polyphaga and survive within cyst walls. Applied and Environmental Microbiology 64:2256-2261.
Stevenson-Hamilton, J. 1911. The relation between game and tsetse flies. Bulletin of Entomological Research 2:113-118.
Su, C., D. Evans, R. H. Cole, J. C. Kissinger, J. W. Ajioka, and L. D. Sibley. 2003. Recent expansion of Toxoplasma through enhanced oral transmission. Science 299:414-416.
Sukanahaketu, S. 1977. The presence of Dientamoeba fragilis in the Ascaris Iumbricoides ova: The first report from Thailand. Journal of the Medical Association of Thailand 60:256-258.
Telford, S. R., Jr. 1994. Plasmodia of reptiles.
Pp. 1-71 in J. P. Kreier (editor), Parasitic protozoa, Vol. 7. Academic Press, San Diego, CA.
Telford, S. R., Jr. 1995. The kinetoplastid hemoflagellates of reptiles. Pp. 161-223 in J. P. Kreier (editor), Parasitic protozoa, Vol 10. Academic Press, San Diego, CA.
Tezcan-Merdol, D., M. Ljungstrom, J. Winiecka- Krusnell, E. Linder, L. Engstrand, and M. Rhen. 2004. Uptake and replication of Salmonella enterica in Acanthamoeba rhysodes. Applied and Environmental Microbiology 70:3706-3714.
Todd, K. S., and D. M. Hammond. 1971. Coccidia of Anseriformes, Galliformes, and Passeriformes. Pp. 234-281 in J. W. Davis, R. C. Anderson,
L. Karstad, and D. O. Trainer (editors), Infectious and parasitic diseases of wild birds. Iowa State University Press, Ames, IA.
Tyzzer, E. E. 1934. Studies on histomoniasis or “blackhead” infection, in the chicken and turkey. Proceedings of the American Academy of Arts and Science 69:189-264.
Valkiunas, G. 2005. Avian malaria parasites and other haemosporidia. CRC Press, Boca Raton, FL.
Van der Ploeg, L. H. T. 1987. Control of variant surface antigens switching in trypanosomes. Cell 51:159-161.
van Riper, C., III, C. T. Atkinson, and T. M. Seed.
1994. Plasmodia of birds. Pp. 73-140 in J. P. Kreier (editor), Parasitic protozoa, Vol. 7. Academic Press, San Diego, CA.
van Riper, C. H., III, S. G. van Riper, M. L. Goff, and
M. Laird. 1986. The epizootiology and ecological significance of malaria in Hawaiian land birds. Ecological Monographs 56:327-344.
Visvesvara, G. S. 1999. Pathogenic and opportunistic free-living amebae. Pp. 1383-1390 in P. R. Murray,
E. J. Baron, M. A. Pfaller, F. C. Tenover, and R. H. Yolken (editors), Manual of clinical microbiology. 7th ed. ASM Press, Washington, DC.
Visvesvara, G. S., and J. K. Stehr-Green. 1990. Epidemiology of free-living ameba infections. Journal of Protozoology 37^5S-33S.
Volkman, S. K., and D. L. Hartl. 2003. A game of cat and mouth. Science 299:353-354.
Warner, R. E. 1968. The role of introduced diseases in the extinction of the endemic Hawaiian avifauna. The Condor 70:101-120.
Wells, E. A., and W. H. R. Lumsden. 1969. Trypanosome infections of wild mammals in relation to trypanosome diseases of man and his domestic stock. Pp. 135-145 in A. McDiarmid (editor), Diseases in free-living wild animals. Symposia of the Zoological Society of London No. 24. Academic Press, London, UK.
Wilson, M. R., and J. B. McAuley. 1999. Toxoplasma. Pp. 1374-1382 in P. R. Murray,
E. J. Baron, M. A. Pfaller, F. C. Tenover, and R. H. Yolken (editors), Manual of clinical microbiology. ASM Press, Washington, DC.
Wilson, V. J. 1972. Observations on the effect of dieldrin on wildlife during tsetse fly Glossina morsitans control operations in eastern Zambia. Arnoldia (Rhodesia) 5:1-12.
Woodworth, B. L., C. T. Atkinson, D. A. LaPointe,
P. J. Hart, C. S. Spiegel, E. J. Tweed,
C. Henneman, J. LeBrun, T. Denette, R. DeMots, K. L. Kozar, D. Triglia, D. Lease, A. Gregor,
T. Smith, and D. Duffy. 2005. Host population persistence in the face of introduced vector-borne diseases: Hawaii amakihi and avian malaria.
Proceedings of the National Academy of Sciences of the United States of America 102:1531-1536.
Yilmaz, S. M., and S. H. Hopkins. 1972. Effects of different conditions on duration and infectivity of Toxoplasma gondii oocysts. Journal of Parasitology 58:939-939.
More on the topic LIFE HISTORY TYPES AMONG PROTOZOA:
- LIFE CYCLE STRATEGIES AMONG PROTOZOA
- Hamsters are host to numerous enteric protozoa, which are often listed and discussed in reviews of parasites, but very few protozoa have any pathogenic significance in the hamster.
- 7 Life History
- INTRODUCTION TO PROTOZOA
- Life History Diversity
- Life History Continua
- The Evolutionary History of Life
- There are trade-offs between current reproduction and other life history traits
- Other Enteric Protozoa
- CONCEPT 7.1 Life history patterns vary within and among species.
- Mode of reproduction is a basic life history trait
- CONCEPT 7.2 There are trade-offs between life history traits.
- CONCEPT 7.4 Life history patterns can be classified along several continua.
- Life tables can be based on age, size, or life cycle stage
- Vaughn Marc M.. The History of Ukraine and Russia: The Tangled History That Led to Crisis. History Demystified,2022. — 164 p., 2022
- The history of HIV is brief but dense. It is filled with aggressive debates, politics, human tragedy, a rich vein of research in medical science, and a dramatic change in drugs and tests in 1996 that translated immediately into better health and longer life.
- Birth is a water-shed event in human life, necessitating various physiological changes to adapt from intrauterine to extrauterine life.
- Types of Water Pollutants
- Types of Environment