MAP Antigens
The vast majority of MAP protein studies in the literature are diagnostic-focused. This focus is well placed given the tremendous challenge in diagnosing paratuberculosis and the need for improved tests that may encourage national control efforts and inform the animal producer on management strategies.
Two of the most prominent challenges involve false positive results and low sensitivity. Sera from animals exposed to environmental or ubiquitous mycobacteria, such as M. avium, will cross-react with MAP antigens, giving a false positive reaction. ELISA test sensitivities have lagged behind specificity making detection of early infection nearly impossible. This may be due to the disease characteristics, which are marked by a low antibody response early after infection such that diagnostic sensitivities in pre-clinical stages will likely be low. Herd-level testing with the goal of national control of paratuberculosis recently estimated the serum ELISA as at least 60% sensitive and 100% specific among Irish dairy herds (Sergeant et al., 2019). Although this sensitivity value has improved over the years, the search for an antigen that can be used to detect infection before disease signs appear would be of considerable value. Two commercial ELISAs used for another chronic disease of small ruminants caused by Mycoplasma agalactiae show test sensitivities of 84%, while specificity was 95.7% for a wholecell antigen with slightly lower sensitivity for a recombinant antigen ELISA (Poumarat et al.,2012). However, these test values were obtained using well-defined serum samples, rather than field sampling dairy herds. Furthermore, 9 of 10 commercially available ELISA tests used to detect the bovine parasitic disease, neosporosis, are all above 95% sensitivity and specificity (Alvarez- Garcia et al., 2013). Examples from these animal diseases suggests that MAP ELISA tests could be improved dramatically.
Is MAP really that immunologically stealthy? Especially as it relates to an antibody response? Perhaps not as sets of three or four recombinant MAP proteins can yield increased ELISA sensitivities (>85%) and specificities (9 7%) (Leroy et al., 2007; Li et al., 2017a), but these are not commercially available yet. Thus, MAP serological detection via current paratuberculosis ELISA tests has set a low bar for researchers to jump over. There are some promising new MAP antigens discussed below that may be developed into a commercial test.8.3.1 Multi-antigen studies
Historically, researchers have investigated single MAP antigens using unclear rationale for why that particular antigen was selected for analysis. Furthermore, these antigens were not benchmarked against other MAP proteins or a commercial ELISA test. However, more benchmarked multi-antigen studies are now being reported. This more comprehensive method was needed and hence systematic approaches to screen all the antigens produced by MAP were conducted. Prior to genome sequencing, this was done by screening an expression library constructed in Escherichia coli (Bannantine and Stabel, 2001; Willemsen et al., 2006). Here the E. coli host is forced to express randomly sheared DNA fragments from MAP. Provided the expression library contains all of the DNA fragments that constitute the entire genome, this expression library should theoretically represent the entire proteome. However, many of the cloned fragments are not present in the correct orientation or are not in frame with the promoter designed to express the cloned fragment. Moreover, E. coli expresses MAP proteins at widely varying levels, many of which are not expressed at all due to toxicity (Bannantine et al., 2010). None the less, this system yielded some solid antigen candidates. Most notable was the study by Willemsen et al. (2006), which identified MAP_2609 and MAP_2942c by phage library screening with serum from a clinical cow. These same antigens were later identified via a protein array/ELISA combination screen with serum from cows at various stages of paratuberculosis (Bannantine et al., 2017a; Li et al., 2017a).
More recently, the M. tuberculosis protein array has been used as a screening tool to identify antigens diagnostic for paratuberculosis (Bannantine et al., 2017a). Although not as closely related to MAP as members of the MAC complex, M. tuberculosis still has over half of its proteins with greater than 75% identity to MAP. Candidate antigens were further tested by ELISA using well-characterized serum samples that encompass all paratuberculosis stages. Rv2878c (MAP_2942c), a disulfide oxidoreductase, emerged as the strongest antigen during clinical disease. However, 10 M. tuberculosis antigens were identified that appear to be detected at early stages of paratuberculosis. This pilot study was then expanded to test 180 well-characterized serum samples from cows in all disease stages (Li et al., 2017a), adding more significantly reactive antigen candidates while confirming those discovered in the pilot study (Bannantine et al., 2017a). Interestingly, the Li et al. (2017a) study also revealed 27 antigens detected in early disease cows that were faecal culture-negative and ELISA negative but had high exposure to MAP, suggestive of the animal being infected, but not yet shedding. This result increases the possibility of developing an ELISA test with the needed sensitivity to detect early MAP infection in a dairy herd.
Another 2-D sodium dodecyl sulfate-po- lyacrylamide gel electrophoresis (SDS-PAGE) separation and immunoblot study with sera from control and infected cattle was conducted to identify proteins reactive only with the infected group. Six proteins were excised from the 2-D gels and identified by mass spectrometry including Hsp65 and MMP (MAP_2121 c) (Piras et al., 2015).
Other formats, such as a multiplex beadbased immunoassay, have been used on serum and milk samples. Beads were coated with six antigens identified from the Li et al. (2017a) study and used to test 180 serum and 90 milk samples in this bead assay format (Li et al., 2017b). The results showed that the multiplex assay has higher sensitivity and specificity than the same proteins used in an ELISA format.
Also, when using milk samples, areas under the curve (AUCs) of the recombinant protein multiplex assay were higher compared with the commercial IDEXX ELISA (Li et al., 2017b). Combining these antigens helped increase the sensitivity further, but then specificity was reduced. These results are promising and suggest improvements to antigen-based tests for paratuberculosis are on the horizon.8.3.2 Johnin purified protein derivative
The MAP purified protein derivative (PPD), also known as Johnin, is a complex secreted protein preparation that appears to vary greatly between preparations. Therefore, this antigen is very difficult to standardize for diagnostic use. It is used in two diagnostic tests, the single intradermal skin test to assay for delay-type hypersensitivity and the IFN-γ test. Both tests measure a component of the cell-mediated immune response. Although this has been considered a great antigen for these tests, its lack of standardization resulted in widely variable test results. Each PPD batch needs be checked for potency, which is performed using a 72-h guinea pig assay using a method comparable with that for tuberculin (Frankena et al., 2018). In an effort to develop a reproducible reagent for these tests, proteomic analyses by 2-D SDS- PAGE were conducted on MAP culture filtrates and 125 proteins were initially identified (Leroy et al., 2007). More recently, Wynne et al. (2012) characterized five PPD preps and showed that while the proteins present in each prep were remarkably consistent, they varied in terms of abundance within each prep. Time of culture harvest, ranging from 20-34 weeks, also had a significant influence on which proteins were present in the final PPD. Finally, a few proteins were identified that were consistently more abundant in the Johnin PPD preps when compared with the avium PPD preps. In separate studies, 194 total PPD proteins were identified with only 10 shared among all three preps analysed (Capsel et al., 2016) and 156 PPD proteins were mentioned by Santema et al.
(2009), but only 25 were listed in that publication. In comparing the proteins identified among these three proteomic PPD studies (Santema et al., 2009; Wynne et al., 2012; Capsel et al., 2016), it was discovered that a core of 15 MAP proteins were identified from all three studies (Fig. 8.1, details in Table 8.2). These 15 proteins likely represent those in highest abundance in PPD extracts. Although these proteins are theoretically secreted based on the way the preps are made, these 15 proteins include membrane, cytoplasmic and periplasmic proteins, as well as those that could not be predicted by the PSORTb software (Yu et al., 2010). It is likely that PPD comprises at least 300 proteins, thus we still have a long way to go in characterizing this useful, but hard to prepare antigen.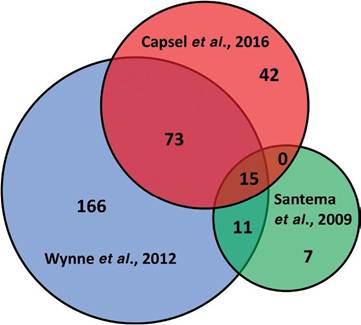
Fig. 8.1. Venn diagram of Mycobacteria avium subsp. paratuberculosis (MAP) proteins in three published studies of MAP purified protein derivative. (From Santema et al., 2009; Wynne et al., 2012 and Capsel et al., 2016.)
8.3.3 Antigens tested in the IFN-γ assay
Cattle develop a proinflammatory T helper 1 (Thl) response to MAP early in the course of paratuberculosis. IFN-γ release assays measure T-cell production of IFN-γ in response to antigen exposure thus providing an assessment of cell- mediated immunity. Measurement of IFN-γ production has shown promise for paratuberculosis diagnosis at early stages of infection and is an indicator of the Th1 immune response, but the specificity is low due to extensive crossreactions with closely related mycobacteria. Whole blood from naturally infected cows with subclinical disease shows significantly higher IFN-γ secretion than blood from healthy control cows after stimulation with the T-cell mitogen, concanavalin A, or a whole-cell extract of MAP (Stabel, 1996). Historically, MAP PPDs have been used as the stimulating antigen for the paratuberculosis IFN-γ assay.
A MAP wholecell extract has been compared with PPDs, but PPDs appeared to be a stronger IFN-γ stimulator (Robbe-Austerman et al., 2006). Five mycobacterial lipids have also been tested this way and none can detect MAP-infected goats (Souriau et al., 2017) or sheep (Thirunavukkarasu et al.,2013). Conversely, young calves and uninfected cows often respond to PPDs used in the IFN-γ assay without having any evidence of infection (Mcdonald et al., 1999) and there exist wildly fluctuating results when the same animals are serially tested (Huda et al., 2003; Dernivoix et al., 2017; Souriau et al., 2017). Another limitation of this assay is the need for fresh whole blood or peripheral blood mononuclear cells (PBMCs) to conduct the antigen stimulation. Samples that have been kept for longer than 12 h show a pronounced IFN-γ decrease in response to antigens (Robbe-Austerman et al., 2006). Some have tried to lengthen the usefulness of the samples out to 24 h before starting the antigen stimulation (Buza et al., 2004; Jungersen et al., 2012; Mikkelsen et al., 2012). Not all of these difficulties can be removed simply by changing the antigen, but the goal of improved early detection and increased specificity has motivated researchers to test purified recombinant antigens in an effort to circumvent the problems associated with PPDs and whole-cell extracts.
Table 8.2. Mycobacteria avium subsp. paratuberculosis (MAP) proteins common to three proteomic purified protein derivative (PPD) studies.
| Locus tag | Gene | aa length | PSORTb location | Description |
| MAP_0151 c | - | 151 | Unknown | Transcriptional regulator TetR |
| MAP_0494 | - | 318 | Unknown | Putative oxidoreductase |
| MAP_1138 c | lprG | bgcolor=white>239Membrane | Lipoprotein | |
| MAP_1339 | - | 148 | Unknown | Hypothetical protein |
| MAP_1595 | bfrA | 160 | Cytoplasmic | Bacterioferritin |
| MAP_1609 c | fbpB | 331 | Unknown | Antigen 85B, mycolyltransferase |
| MAP_1889 c | wag31 | 261 | Cytoplasmic | Antigen 84 |
| MAP_1997 | acpM | 116 | Cytoplasmic | Acyl carrier protein |
| MAP_2451 c | atpD | 403 | Membrane | ATP synthase [beta] chain |
| MAP_2677 c | - | 133 | Unknown | Putative oxidoreductase |
| MAP_3362 c | sahH | 497 | Cytoplasmic | Adenosylhomocysteinase |
| MAP_3531 c | fbpC2 | 353 | Periplasmic | Antigen 85C, mycolytransferase |
| MAP_3840 | dnaK | 624 | Cytoplasmic | 70 kD heat shock protein |
| MAP_3936 | groEL2 | 542 | Cytoplasmic | 60 kD chaperonin 2 |
| MAP_4143 | tuf | 397 | Cytoplasmic | Elongation factor EF-Tu |
Recently, six recombinant antigens have been examined in the IFN-γ assay using a 20h PBMC stimulation from animals thoroughly characterized with a PPD-IFN-γ assay at the study's outset (Dernivoix et al., 2017). None of these recombinant proteins showed higher sensitivity than the PPD, although three proteins were able to distinguish MAP-infected from M. bovis-infected animals. In a separate study, the stress-induced proteins, MAP_2698c and MAP_3567, were examined by this format for sheep vaccinated with the same antigens. A 48h stimulation resulted in IFN-γ responses, but varied depending on the adjuvant used (Gurung et al., 2014b). Two other recombinant proteins, MAP_0268 c and MAP_3651 c, have been used to stimulate IFN-γ responses in sheep (Hughes et al., 2013) and cattle (Hughes et al., 2017) and have shown promise in both. Another study examined 16 recombinant proteins in goats (Souriau et al., 2017). The proteins encoded by MAP_3651 c and MAP_1050c emerged as the most promising candidates for detecting infected shedding goats but could not detect infected non-shedding goats. Also, use of these recombinant antigens was discouraged in adult goats (>2 years old) as infection predictions dropped below acceptable levels. A final study examined 15 recombinant proteins in a variety of time and blood preservation conditions (Mikkelsen et al., 2012). Thus, only 41 antigens have been evaluated in this way these past 10 years and none appears promising enough for commercial development. Perhaps a high-throughput method should be engineered to examine more recombinant antigens for this assay.
In general, it appears that good recombinant antigen candidates for serological detection of MAP infection are not good candidates for IFN-γ stimulation assays. Several recombinant proteins have been tested in both assays (MAP_1589 c AhpC, for example), and thus far none works well in both. Also, those that appear to be good candidates in the IFN-γ assay are not listed among the good serological antigens and vice versa. Furthermore, it appears that recombinant proteins still cannot stimulate IFN-γ production in PBMCs from infected cows better than PPDs and this has been the main problem with using recombinant proteins in this assay. They aren't as strong an antigen as the PPDs even though they would be easier and more consistent to prepare. As mentioned in Section
8.3.2 PPDs contain hundreds of proteins that stimulate a wide variety of T cells, which a single, or even a dozen, recombinant proteins cannot. Furthermore, many studies have reported high variability in recombinant protein IFN-γ assays for paratuberculosis. This is partly due to the age of the animal, and partly the need for fresh PBMCs to conduct the assay and the differences in time used to stimulate the PBMCs, which can vary from 16-72 h. These obstacles do not mean researchers should stop looking for a strong antigen to incorporate in a marketable paratuberculosis IFN-γ assay, but the approach must be clever and antigen solid before investing too much in this line of research.
8.3.4 Cell-mediated immune (CMI) stimulating antigens of MAP
An early diagnostic antigen will more likely generate a measurable cell-mediated immune response than a humoral response based on what we know about progression of immune parameters to this disease. Several studies have looked beyond the proinflammatory cytokine, IFN-γ, as just one measure of a CMI response. IFN-γ is easy to measure, hence the focus on this assay for early MAP detection. However, measuring any type of cellular-based immunity accurately is difficult due to biological variation, standardization and technical complexity, which is why the MAP IFN-γ assay itself has shown high variability. To characterize a CMI response, the circulating immune cells and their subsets should be quantified by flow cytometry. This approach can identify dendritic cell maturation and activation, which drives the Th1 response. The skin test measures delayed type hypersensitivity through antigen-specific memory T cells, which creates a local inflammation at the injection site from recruitment of other immune cells. T-cell activation can be measured either by changes in surface markers (CD25, CD45RO, CD69) or by measuring intracellular cytokine levels. MAP proteins have played a role in all of these CMI responses.
While MAP itself has been shown to activate resident dendritic cells (DCs) many times, MAP effector proteins that cause DC activation have also been recently discovered. Antigens with this capability are well positioned as good vaccine candidates since DCs provide an important link between innate and adaptive immunity. MAP_1948 (Byun et al., 2012), MAP_1981c (Kim et al., 2018b), MAP_2541c (Kim et al., 2018a), MAP_1305 (Lee et al., 2014) and MAP_1569 (Lee et al., 2009; Noh et al., 2012) have all been shown to promote DC activation. Most of these same proteins have been shown to selectively activate DC through TLR-4. Dendritic maturation can go on to promote cytotoxic lymphocyte killing. Both MAP_2121 c and MAP_1569 promote cytotoxic T cell activity (Noh et al., 2012; Abdellrazeq et al., 2018). Many other studies have examined how the intact Mycobacterium promotes CMI responses, but those are beyond the scope of this antigen- proteomic-focused chapter.
8.3.5 Antigens tested by ELISA
Milk has become a popular sample in recent years with the development of milk ELISA testing, but defined antigens have yet to be tested on milk samples except for one study (Li et al., 2017b). Recently, a sandwich ELISA was developed with a monoclonal antibody to the Apa protein (MAP_1569) and used on faecal eluates to detect that antigen in paratuberculosis-positive cows (de Souza et al., 2018), and this type of approach has been used to measure MAP-specific IgA in the faeces before (Begg et al., 2015). But the most common sample for ELISA historically is serum.
The search for defined antigens to incorporate into an ELISA test began in 1991 with the 85C antigen and a bacterioferritin biochemically purified from a MAP culture filtrate (Sugden et al., 1991). These antigens each showed 86 and 100% sensitivity, with the 400-kDa bacterioferritin antigen detecting all 22 infected animals in that study. Lipid antigens were readily purified and the ELISA format tested with some degree of success as well. Then molecular biology was applied to MAP and recombinant proteins were expressed in E. coli and tested.
Among MAP recombinant antigens incorporated into ELISA within the past 10 years, PtpA (encoded by MAP_1985), has been studied extensively. This protein is a tyrosine phosphatase that is secreted early during infection of the host. It is reported to be antigenic in humans (Slavin et al., 2018), sheep (Gurung et al., 2014a), and displayed a 90% ELISA sensitivity and 91% specificity in cattle (Bach et al., 2018). The Bach et al. (2018) study also showed that the PtpA ELISA outperformed a commercial ELISA for early detection of MAP antibodies in a 55-animal study. Once validated, this could make PtpA the best recombinant antigen for use in a paratuberculosis ELISA test for cattle. PtpA was not among the 729 M. tuberculosis proteins that reacted to sera from paratuberculosis- infected cows, but it should be noted that only 81.6% identity exists between MAP PtpA and M. tuberculosis PtpA (Bannantine et al., 2017a).
Fourteen culture filtrate proteins were shown to be immunogenic for paratuberculosis cows. Five of these were tested in an ELISA format based on strong reactivity with sera from MAP-immunized rabbits (Chaubey et al., 2018). These five include MAP_1693c, MAP2168c, MAP_1569, MAP_352 7 (PepA) and the aforementioned antigen 85C (MAP_3531 c). They were mixed and used as a cocktail for coating ELISA plates and directly compared with a MAP whole-cell extract ELISA.
8.4
More on the topic MAP Antigens:
- Proteins and Antigens of Mycobacterium avium subsp. paratuberculosis
- Study of MAP Proteins
- MAP Immune Responses in Crohn’s Disease Patients
- Genomic Epidemiology of MAP
- Enumeration of MAP
- Cultural Requirements of Different Strains of MAP
- Survival of MAP During Dairy Processing
- Comparison of the Virulence and Pathogenicity of MAP Strains
- Innate Response to MAP Infection
- Genomic Comparison of MAP Strains
- General Principles for the Cultivation of MAP