RABBIT HAEMORRHAGIC DISEASE
J. PAUL DUFF
A nimal Health and Veterinary Laboratories Agency Diseases of Wildlife Scheme (AHVLA DoWS), Great Britain Wildlife Disease Surveillance Partnership, Penrith, Cumbria, UK Rabbit haemorrhagic disease (RHD) (synonyms viral haemorrhagic disease (VHD), necrotic hepatitis, rabbit plague, rabbit X disease, rabbit calicivirus disease) is a highly contagious and infectious, acute but occasionally subacute, disease caused by the RHD virus (RHDV), a calicivirus.
The only species affected is the European rabbit (Oryctolagus cuniculus).AETIOLOGY
Initially RHD viral identity was unclear but a calicivirus aetiology is now accepted and supported by virus sequencing. RHDV belongs to the genus Lagovirus, which is used generically to refer to caliciviruses of lagomorphs. RHDV is antigenically related to, but distinct from, the calicivirus that causes European brown hare syndrome (EBHS), with which it shares approximately 71% nucleotide identity. RHDV is not closely related to the other known calicivi- ruses. RHDV is a small, round, 32—35 nm RNA virus with typical calicivirus structure and characteristic appearance in electron micrographs. The genome has a single positivesense RNA strand. RHDV has a major capsid polypeptide (60 Kda). Various RHDV strains from around the world show 89—100% identity and with a few exceptions can be considered as one serotype. There has been no observed attenuation (reduction in viral virulence induced by passage through rabbits). The virus provides an opportunity to study viral evolution and viral acquisition of pathogenic properties. RHDV has quasi-species characteristics — that is, an array of slightly different RNA viruses, existing as many related but distinct ‘strains’ that are immunologically cross-protective to some degree. The complex inter-relationship between these is important in the understanding of the pathogenesis and epidemiology of the disease.
A selection of strains distinct from the diseasecausing strain require a brief mention.Infectious Diseases of Wild Mammals and Birds in Europe, First Edition. Edited by Dolores Gavier-Widen, J. Paul Duff, and Anna Meredith. © 2012 Blackwell Publishing Ltd. Published 2012 by Blackwell Publishing Ltd. Material authored by Paul Duff remains Crown Copyright.
• A smaller, smoother RHDV particle is detectable in some RHD livers and may arise from immunological degradation processes; it is a marker for chronic/subacute disease.
• A rabbit calicivirus related to RHDV was isolated from healthy farmed rabbits in Italy and named rabbit calici virus (RCV). Significantly RCV antibodies are cross- protective to RHDV infection. This virus is not virulent but is present at relatively high seroprevalence and replicates in the intestine rather than the liver. It is similar but not identical to the non-pathogenic RHD-like viruses identified in France and Australia.
• Another RHDV subtype, called RHDVa, causes disease in farmed rabbits but not in properly vaccinated animals. This variant varies from ‘classical RHDV’ in several amino acid substitutions, which alter the main neutralizing surface epitope.
• RHDV-like viruses; several countries including the UK, Czech Republic, Australia and New Zealand reported RHD antibodies from archived sera from animals sampled before 1984. These antibodies provide variable cross- protection to RHDV infection and in some of these populations may be the reason behind the variably low mortality seen following infection with classical RHDV.
Unlike some caliciviruses, RHD shows high species specificity. Experimental infection studies in a range of wild and domesticated bird species and 13 mammalian species(2), including lagomorphs such as the brown hare (Lepus europeaus) have not reproduced the disease. There are no accidental hosts, although foxes and some other natural predator species produce antibody without viral replication.
EPIDEMIOLOGY
Unusually, RHD first emerged as a point disease outbreak in place and time in China in 1984. Here it was seen initially in Angora rabbits imported from Germany a few days after they first arrived in China(3). In less than 9 months it had spread to rabbitries within an area of 50,000 square kilometres, killing 500,000 farmed animals, and quickly spread to Korea. Independently, RHD was reported in domesticated rabbits in Italy in October 1986; whether it entered Italy from China is not known. By 1988, RHD had spread over large areas of the globe, including much of Europe, Russia, the Middle East and parts of Africa, Cuba, Mexico, USA, Uruguay and India, and in each was identified initially in farmed animals and subsequently in wild rabbits where these occurred. For instance, the disease occurred first in pet rabbits in southern England in 1992 (investigations could not find a likely source), and wild rabbits there in 1994, and by 1996 the disease was relatively widespread in both farmed and wild rabbits throughout Britain and Ireland.
Possible sources of RHDV include: mutation from the EBHS virus; mutation from a pre-existing non-pathogenic virus of rabbits; or transfer of a hitherto unknown virus from another animal species(2). Complimentary serology results indicate that the second possibility is the more likely. Emergence of pathogenic virus may have occurred, or recurred, on several occasions. There may be a repeating cycle of evolutionary change whereby non-pathogenic, finely host-adapted strains change, probably by mutation, to highly pathogenic strains, and then subsequently change back by mutation to non-pathogenic forms.
The epidemiology of RHD is not well understood and the occurrence of outbreaks cannot be reliably predicted. Several factors are involved at the rabbit population level and include the interplay of the range of virus strains coupled with the presence or absence of respective neutralizing (protecting) antibody populations.
Other important epidemiological factors include the presence of protective antibody in metapopulations before the arrival of clinical RHD; the lack of susceptibility of very young rabbits independent of their immunological status; the interplay with myxomatosis; the timing of breeding; and finally, seasonality.RHD can be expected wherever there are wild rabbits in Europe. The disease has been found in isolated populations, particularly those on offshore islands. Maternal immunity is protective to about 6, or exceptionally 8, weeks of age. By some unknown mechanism not related to humoral immunity, young rabbits up to about 4—6 weeks of age are not susceptible to disease, and this allows a window of opportunity whereby exposure to the virus allows kits, in the absence of maternal antibody, to develop a lasting immunity1-4). In effect, the innate protection ensures that rabbits in this age class will survive, and this period of juvenile protection therefore reduces the epidemic intensity and enhances subsequent population recovery.
Male and female rabbits are equally affected, and pyrexic females may abort prior to death. The disease in Europe shows variable seasonality and may occur in any month; however, in England it is more frequent in the autumn, whereas in most of France and Spain there is little seasonality. In Australia and parts of Spain rabbit populations in the drier areas appear to suffer long-term population reductions more than those in wetter regions. Vectors are important and wild carnivores and scavenger species, particularly gulls, are an important mechanical vector of infection. Mechanical transport of virus probably explains the stepwise progression of new epidemic fronts, for example with leaps of up to 280 km in England. Insect transmission exists and is the probable reason behind the escape of the virus in 1995 from the Wardang quarantine island, 5 km off the Australian coast, to the mainland and subsequent spread in Southern Australia.
Windborne insect transmission from continental Europe may have accounted for an isolated outbreak of RHD on the east coast of England. There are no known reservoir animal species and it would be reasonable to assume that the virus would be removed from a population after it had killed all susceptible animals (fade- out). Viral RNA can be detected in populations after mortality ceases but does not necessarily equate to rabbit carrier status, as RNA may not be infective virus. Virus persistence in the environment occurs — for example, burrow systems may act as environmental reservoirs and rabbits themselves may be passive vectors through fur contamination. Clinical RHD may become almost endemic, persisting for years, owing to the continual circulation of RHDV in some large, heavily infected populations. Reintroduction of virus by the means listed above and by human actions regularly brings virus back into populations. Molecular evidence indicates that RHDV has been introduced to many countries on several occasions often by human activity, e.g. in moving subclinically infected live rabbits and rabbit meat. The rapid replacement of immune rabbits by susceptible animals is facilitated by the high natural mortality of rabbits (60% per annum). Mortality is difficult to assess and significant numbers of affected animals die undetected in warrens underground. In Australia RHD caused population reductions of 85% in arid areas, and 73% in coastal areas in year one, decreasing to a 12% reduction by the third year. However, the impact of RHD has declined in many populations over the years since first infection. High- intensity outbreaks in Spain suggested the existence of complete population immunological naivety, whereas the variable intensity in other countries indicated pre-existing antibody protection.In Spain RHD can cause two different pictures in adjacent rabbit colonies: one may crash, whereas the other has low mortality. In this country in particular, rabbit population recovery has been variable.
The greatest losses in Spanish colonies occurred in areas where rabbits were already in low numbers1-5); when epidemic disease occurred it frequently recurred to the detriment of population recovery. A similar but less noticeable impact was seen in UK populations, where those experiencing heavy initial reductions due to RHD showed a poor recovery whereas populations with low initial mortality recovered well. Initial mortality on Ramsey, a UK marine island, was estimated at 5,000 deaths, with 40 deaths/ha. The outbreak was still active 4 months after onset. Outbreaks recurred on the island for three consecutive years and the population declined by 50% between 1994 and 1998. Further epidemiological postulations and questions that may be relevant, but are difficult to prove, include the following.• RHDV has circulated in an avirulent form in Britain and possibly Europe for at least 50 years(6).
• Why avirulent RHDV changed to a pathogenic phenotype, what stimulated the change and whether the change was entirely due to viral mutation are not known.
• Serologically naive rabbits may be infected with RHDV, or RHDV-like non-pathogenic virus, and not show overt disease.
RHD has not had a significant impact on the national wild rabbit population of several northern European countries, including the UK. Over recent years more RHD-like viruses have been identified. These have significance for understanding genome composition, molecular epizootiology and evolution of pathogenicity(7). It remains possible that the virulence of RHDV for rabbits may not be defined simply in terms of unique genetic determinants of the viral genotype.
TRANSMISSION
I nfection is by direct transmission, and indirect transmission facilitated by the stability of the virus in the environment. Excretions and secretions are infective, as are carcasses and fomites such as bedding material. Infection can occur by oral, nasal, conjunctival and parenteral (insect bite) routes. Viral RNA is present in urine and faeces for up to 4 weeks post-infection in surviving animals. Virus persists well in the environment for up to 1 month at least; viral activity is not reduced by pH 3.0 conditions, and it persists experimentally in rabbit tissues for 18 months and for 100 days in dry cloth. It is stable with freezing (e.g. in frozen rabbit meat). Polymerase chain reaction (PCR) studies have shown RHDV nucleic acid in wild rabbit bones 7 years after an outbreak. Spread of virus in fomites and spread by people (on tools, equipment and vehicles) contaminated with virus is important in disease dissemination. Haematophagous insects, fleas and mosquitoes spread virus mechanically, while the oral and faecal excretions of flies (fly spots) are infectious. Transmission by flying insects has been considered as a reason for epidemics of the disease to ‘jump’ by large distances ahead of the disease front, disrupting any epidemic front. Spread by scavenger species, particularly birds, over long distances and onto islands also occurs and is important. Death is rapid, and consequently the period of clinical infectivity in the individual is short. Predators, including primary rabbit predators such as foxes and eagles, may excrete infective virus unchanged by digestive processes for short periods in their faeces, thereby potentially facilitating long-distance viral dissemination.
PATHOGENESIS, PATHOLOGY AND IMMUNITY
The incubation period is 1—3 days and death usually occurs 12—36 hours after onset of clinical signs. The clinical disease can be peracute, acute, subacute or, rarely, chronic. Infection only occurs in rabbits older than 40—50 days; the mechanism for this resistance is not known, but humoral immunity is not involved. Possibly implicated is a combination of lack of viral receptors in cells of the digestive and respiratory tracts and the innate immune physiology of the neonate.
Following entry of the virus into the rabbit body and infection through the mucosae of the respiratory and digestive tracts, there is limited local spread, primary replication and then haematogenous spread to the principal target organ, the liver. Replication of the virus in hepatocytes leads to cell death and apoptosis, presumably with
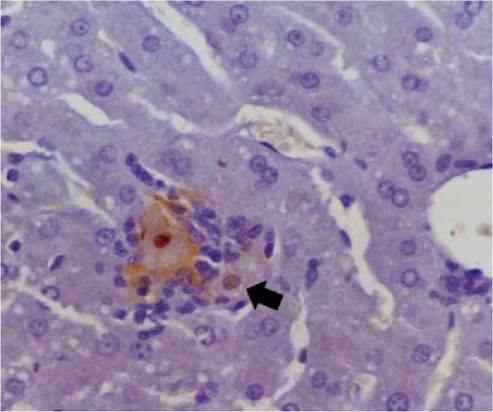
FIGURE 5.1 Histological section of liver of a rabbit 24 hours after oral inoculation with RHDV. A single hepatocyte is surrounded by inflammatory cells. Viral antigen (reddish brown coloration) is within the nucleus and on the cell membrane of this hepatocyte. Note the presence of antigen in the nucleus of an adjacent hepatocyte (arrow). Immunoperoxidase with primary monoclonal antibody to RVHD- virus. Photo: D. Gavier-Widen, SVA.
local spread to neighbouring liver cells (Figure 5.1) and further hepatocyte necrosis (Figure 5.2), and ultimately to liver failure. The virus may also be isolated from the spleen (Figure 5.3), and occasionally from other organs. Most rabbits develop terminal disseminated intravascular coagulation (DIC) and shock. Occasional abortions are probably related to pyrexia, as viral infection of placental tissue and fetal tissue have not been demonstrated.
Humoral antibodies are frequently produced despite the rapid progress of clinical disease (antibodies are even detectable at 5—7 days post-infection, A. Lavazza, personal communication). Young rabbits with innate resistance to RHDV may develop immunity on exposure. It is assumed that surviving rabbits have a long, possibly life-long, immunity to RHDV. It is assumed that ‘fitness’, as assessed by other methods, offers no effective protection to RHD infection.
As the course of the disease is rapid, animals found dead are usually in good condition. RHD gross pathology is variable, lesions may be subtle and there are no consistent characteristic findings, belying the acute nature of the disease and extensive liver necrosis that causes death. In peracute disease no obvious gross lesions may be apparent.
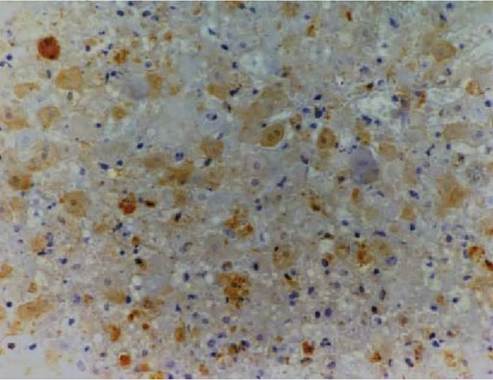
FIGURE 5.2 Histological section of liver of a rabbit that died 48 hours after oral inoculation with rabbit haemorrhagic disease virus. Viral antigen (brown coloration) is within the nucleus and cytoplasm of hepatocytes and along their cell membranes, in necrotic hepatocytes, debris and in a macrophage. Immunoperoxidase with primary monoclonal antibody to RHDV. Photo: D. Gavier-Widen, SVA.
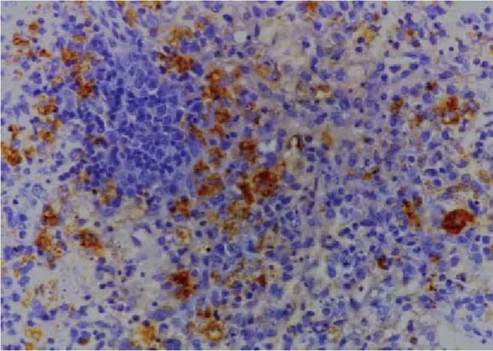
FIGURE 5.3 Histological section of spleen of a rabbit euthanized 4 days after oral inoculation with RHDV. Viral antigen is in the cytoplasm of macrophages. Immunoperoxidase with primary monoclonal antibody to 60 kD capsid protein of RHDV. Photo: D. Gavier-Widen, SVA.
The acute form is most frequent, with hepatosplenomeg- aly(7). The liver is often a yellowish colour, and lobule necrosis may give rise to a reticular pattern. Haemorrhagic carcasses are relatively infrequent (the disease name is somewhat of a misnomer) and the variable presence of petechial and ecchymotic haemorrhage in various organs, pulmonary oedema and thoracic transudates and the presence of blood without clots have two causes: DIC and coagulopathy. However, haemorrhage, tissue congestion due to DIC and shock are not specific to RHD. Four lesions, although not specific for RHD, are reasonably consistent: i) liver changes as noted; ii) intense, sometimes violaceous, congestion of the tracheal mucosa often associated with copious tracheal froth; iii) marked congestion of the spleen; iv) variable lung changes, which may include haemorrhage, patchy congestion and thoracic transudates. There may also be blood- stained froth and blood clots around the nares and a mild catarrhal enteritis.
Histopathology reveals distinctive changes in the liver characterized by extensive (massive) liver necrosis, congestion and haemorrhages in animals that die peracutely to zonal hepatic necrosis, typically periportal and of variable extension in acute and subacute disease. Hepatocyte destruction may include nuclear pyknosis and karyor- rhexis. Inflammatory infiltrates are moderate and consist of lymphocytes in portal spaces and sinusoids. There may be early calcification in survivors. There may be microthrombi and extensive haemorrhages in the lungs and kidney and lesions in the spleen vary from congestion to lymphoid necrosis associated with lymphopenia. Chronic disease pathology is putative and very infrequent if it does occur, and apart from non-specific mild icteric coloration of blood vessels the rare survivors do not have residual lesions.
CLINICAL SIGNS AND TREATMENT
Disease may be peracute with no clinical signs seen early in the epidemic and animals are found dead; acute (most frequent) and subacute disease show milder signs. Acute infection may produce clinical signs for 12—36 hours, with fever (>40°C), dyspnoea and epistaxis (Figure 5.4A) although bleeding from orifices is not always seen and not pathognomonic. Nervous signs, primarily as a result of hepatic encephalopathy, are frequent and include depression (Figure 5.4B), convulsions, incoordination, fitting, opishthotonos and vocalization (shrill screaming). In large outbreaks some rabbits are subclinically affected, and jaundice is reported in these before death(7).
There is no effective treatment for domesticated or wild animals. Hyperimmune sera may have a role in protection in the face of an outbreak but is probably impractical.

FIGURE 5.4 Wild European rabbits that died during a confirmed RHD outbreak on Ramsey Island, Wales. A. One dead animal shows bleeding from the nose, a clinical feature of some, but not all, RHD cases. B. The other rabbit shows profound depression, lying in abnormal recumbency in the open during daylight. Photo courtesy Roger Trout, Rabbit Wise, Hants, UK.
DIAGNOSIS
For detection of the virus the liver should be examined, as it contains the highest viral titre. In vitro isolation by virus culture has never been achieved. A haemagglutinin test utilizing human erythrocytes (type O) has been used; however, electron microscopy, enzyme-linked immunosorbent assay (ELISA), western blot, PCR and immunohistochemistry (to demonstrate RHD antigen in hepatocytes and splenic histiocytes) are more sensitive tests. Experimental infection of domestic (seronegative) rabbits is not usually necessary but can be employed in cases where diagnosis is equivocal
For serology, the tests most frequently used are haemagglutination inhibition or ELISA. Owing to the acute nature of the disease, early deaths may be caused by high titre viral infection in the absence of detectable antibody, and at the other extreme is the possibility of chronic low viral titre cases in the presence of antibodies (especially IgM). The diagnosis of RHD should be based on a range of observations from clinico-pathology and laboratory tests to histopathology. Clinical history of mass mortality in a rabbit warren affecting adult rabbits but not neonates over a period of up to 6 weeks should arouse suspicions of RHD. Myxomatosis in wild rabbits in Europe may occur concurrently. Histopathology will confirm the diagnosis on the basis ofcharacteristic liver lesions in the majority ofanimals. Laboratory confirmation was based for years on electron microscopy, where massed ranks of calicivirus are seen in typical cases; however, ELISA and western blot methods are now more reliable. PCR should be considered the diagnostic method of choice, and this test can detect virus in tissues from rabbits that are severely decomposed or skeletal. Some caution with PCR results should be exercised, however, as positive livers may indicate viral contamination from other sources, or the animals could have a putative chronic form of disease in which lesions are in remission and death is caused by other causes. Population serology is important for epidemiological analysis1-3). Survivors of outbreaks also show seroprevalence, which is difficult to interpret, although the use of ELISA serology tests to differentiate serotypes is informative.
MANAGEMENT, CONTROL AND REGULATIONS
There is no effective treatment for RHD in domesticated or wild animals. Humane euthanasia should be considered for affected individuals.
There are no current EU regulations for RHD. The disease in domesticated and wild rabbits is reportable to the World Organisation for Animal Health (OIE). RHD was a notifiable disease in both wild and domesticated rabbits in some European countries, as in the UK between 1996 and 1998 but was denotified in the UK (and New Zealand) when it was no longer feasible to contain and eradicate the virus. The virus is generally resistant to many commonly used disinfectants however it is inactivated with 10% sodium hydroxide and 1.2% formaldehyde.
RHD is transmissible between wild and domesticated rabbits by direct and indirect methods, and it is therefore a trade-sensitive disease of importance in the many areas of the world where rabbit farming is commercially significant. RHD in domestic colonies and laboratory populations is controlled by very strict biosecurity and regular vaccination. These methods must be under regular review, as inadvertent introduction of the virus may easily occur.
Vaccination in wild colonies is impractical at present. A simple killed formalin- treated, adjuvanted vaccine is commercially available, providing protection for 6—12 months. Recently, a novel bivalent vaccine utilising a recombinent virus consisting of a laboratory- attenuated myxoma virus strain which has the capsid protein gene of RHDV inserted has become commercially available in Europe, providing 12 months protection against both RHD and myxomatosis1-8). In Spain, rabbits are a valuable species for hunters, and there have been initiatives to develop a recombinant vaccine attached to a live non- pathogenic virus (poxvirus) that would spread vaccine- derived non-pathogenic virus to a limited number of rabbits in contact with the vaccinated individuals. To date, however, this has not been licensed or used in the field. There is some evidence that RHDV strains may persist in regularly vaccinated domestic rabbit populations, although there is no obvious vaccine failure and several factors may be involved in this observation. RHD was eradicated in Mexico, where it was introduced in one shipment of rabbit meat from China; this was probably a result of the absence of wild European rabbits in that country. By contrast, eradication was not possible in Europe when RHD became firmly established in the natural reservoir population)3).
Management of RHD in wild populations globally can have opposing goals: in areas where RHD is used as a means of biological control of pest populations compared with areas in which control of RHD is sought to reverse population declines. Deliberate release of the virus as a form of biological control appears to vary in effect, depending on environmental conditions, time of release, bait quality, host factors, population size and climate. The results of biological control also depend on the presence of non-pathogenic or silent forms of the virus that may be circulating. Humans have had a significant direct and indirect role in transmission of RHDV in wild and domesticated populations in deliberate, identified ways, and covert, or accidental, ways.
PUBLIC HEALTH CONCERN
No other species, including humans, have been reported to be clinically affected by RHDV Therefore the risk to public health from RHD viruses of currently known genomes is considered negligible. There is public concern when RHD mass mortality incidents occur, particularly if bodies are visible, and these frequently prompt requests for diagnosis and health assurance. No evidence has come to light to suggest that the virus crosses species barriers or that it infects people — for example, there was no serological response in 259 people occupationally exposed to the virus.
SIGNIFICANCE AND IMPLICATIONS FOR ANIMAL HEALTH
RHD is a major concern for rabbit breeders globally, and any colony of O. cuniculus no matter how geographically isolated could be infected. In the nearly 30 years since the first report of RHD the disease has caused significant animal losses throughout the world where rabbits are reared (China 140 million deaths, Italy 64 million deaths) and in wild populations. The losses in farmed animals have now been controlled by the regular use of commercial vaccine.
The European rabbit originated in the Mediterranean areas of Southwest Europe, where wild populations are currently in decline. In Spain the decline is considered to be multi-factorial in nature, related to habitat degradation, myxomatosis and RHD; however, since the arrival of RHD in 1988 populations have not recovered)5). This led indirectly to increased hunting pressure on wild predators of rabbits. The combined effects of myxomatosis and RHD are significant in rabbit populations endemically infected with both viruses, but the complex synergy is not fully understood(4). In Spain the rabbit is also obligatory prey for predators, including two critically endangered species, the Iberian lynx (Felis pardina) and the imperial eagle (Aquila adalberti). Conservation of rabbit populations here is the subject of renewed initiatives.
This is contrasted with the ecological picture in Australia, where the rabbit continues to thrive in suitable dry areas of the continent despite the introduction of RHD. Here the rabbit, introduced nearly 200 years ago, still has a deleterious effect on the environment and endemic fauna. By contrast, in Europe, hunters should be
advised to limit their control activities until after outbreak mortality ceases. Translocation of live rabbits and moving rabbit carcasses and viscera is a risk for dissemination of virus and should be avoided, as the effects of the disease introduction to new wild populations are entirely unpredictable. In the UK outbreaks are sporadic with highly variable impact on the rabbit populations, and in a few localities the decline in rabbit population has affected sensitive habitats, e.g. chalk grasslands and their threatened flora and fauna.
There is a potential untested threat from RHD to the world’s rarer lagomorph species.
ACKNOWLEDGEMENT
The help of Dr Antonio Lavazza in several sections is gratefully acknowledged.