EUROPEAN BROWN HARE SYNDROME
DOLORES GAVIER-WIDEN
National Veterinary Institute (SVA), Uppsala, Sweden and Swedish University of Agricultural Sciences
European brown hare syndrome (EBHS), also named leporid caliciviral hepatitis, is a calicivirus infection of free-living and farmed hares characterized by acute hepatitis.
AETIOLOGY
The European brown hare syndrome virus (EBHSV) belongs to the genus Lagovirus, family Caliciviridae. The virus is closely related antigenically to the rabbit haemorrhagic disease virus (RHDV), which causes similar hepatitis in domestic and wild rabbits ( Oryctolagus cuniculi) and with which it shares epitopes of the major capsid protein gene (VP60). No cross- infection between the hare and rabbit disease occurs, probably because of the low homology between the respective EBHSV and RHDV. EBHSV has similar morphology and biochemical properties to RHDV; however, neither can be cultivated i n vitro.
The EBHSV has been cloned and its genome has been fully sequenced. It was shown to be 7442 bases long, to have a nucleotide identity with RHDV of 71% and to have a similar genomic organization1-9). The EBHSV has evolved since the late 1980s, and new isolates have been identified in Europe.
EPIDEMIOLOGY
GEOGRAPHICAL DISTRIBUTION AND HOSTS
EBHSV appears to be highly host-specific, and infection is restricted to hares of the genus Lepus. It affects free-living and farmed European brown hares (Lepus europaeus) and mountain hares (Lepus timidus), and there is one report of EBHS in captive Lepus capensis in Belgium(10). It is not known if other L epus species are susceptible to EBHSV infection. EBHS has not been described in the published peer-reviewed literature in other species. Experimental infection of rabbits with EBHSV results in the development of low levels of antibodies, which do not protect against infection with RHDV.
Similarly, infection of hares with RHDV results in a mild antibody response, which does not protect against EBHSV infection.EBHS was first described as a new epidemic disease of unknown cause in the early 1980s in Sweden. Its viral aetiology was identified in 1988 by electron microscopy of liver homogenates of hares that had died from EBHS, which revealed viral particles that resembled those observed in the liver of rabbits with RHD(11). EBHS occurred in Europe long before the emergence of RHD. Retrospective studies on archived material and old reports showed that European brown hares in England had antibodies to EBHSV as early as the 1960s(12), showed clinical disease and gross pathology consistent with EBHS (but unconfirmed by histopathology and virology) in 1982 and had first confirmation by histopathology in 1990(13). EBHSV was identified by PCR on paraffin sections from the 1970s in Sweden. EBHS occurs today in many European countries, often endemically in free- living hares. EBHS has been reported in Belgium, Denmark, Finland, France, Greece, Slovakia, Sweden, Italy, Germany, UK, Croatia, Austria, Spain, Czech Republic, Poland and Switzerland. EBHS has not been detected outside Europe, except for a report in Argentina. Many seroprevalence studies on hare populations in various European countries have been conducted, showing a wide range in prevalence, from about 11% to up to 95%.
EBHSV is highly contagious and frequently causes short outbreaks of mortality. Mortality rates are high when the outbreaks occur for the first time in the area and the population of hares is immunologically naive. In endemic areas, where the majority of the hares have antibodies, cases are sporadic or mortality rates are low. Further exposure of immune hares to EBHSV results in boosting the antibody response. EBHSV may spread to new areas, often contiguous to endemic areas, causing new mortality events(14). Clustering of cases in areas of more intensive agriculture, where the population density of European brown hares is higher, has been observed1-15).
The time sequence of mortality in free- ranging hares was described in England: between 4 and 50 hares were found dead each week over a 3-week-period. The average number of deaths per incident was 28. In some cases mortality continued for 3—6 weeks(13). In one event, approximately 100 hares died in a period of several weeks; this was estimated to be 60% of the population. More incidents were observed in areas with high population density(13).
Longitudinal studies in various regions of Europe have described seasonal clustering of EBHS mortality, but the seasons are different and cases of EBHS may occur all year round. For example, mortality was observed to be highest in the late autumn (October, November and December) in Sweden, coinciding with the period of highest hare population density in the year, and with a high proportion of hares’ young-of-the-year, which have reached the age of susceptibility for infection1-15) (see host factors below). Similar higher EBHS mortality during the autumn has been observed in England, France, Italy and Denmark. Highest mortality was observed in mountain hares in Finland in the spring and summer(14).
No wildlife reservoirs of EBHSV are known. It has been speculated that European brown hares that survive the infection act as reservoirs. However, maintenance of EBHSV endemically has also been recorded in populations of mountain hares in areas where brown hares are scarce(14). In EBHS endemic areas, a small proportion of healthy hares are subclinical virus carriers, for example in one study in Italy, this was shown to be about 1%, in a population with a seroprevalence 67% and 78% in two consecutive years(16), and in a study in Poland it was 7.6% in a population with a seroprevalence of 38%(17).
EBHS is an important disease of farmed hares in Europe. The mortality varies from 10% to 100%. Mortality is high in the initial outbreak on farms, but often only about 10% of the hares succumb to EBHS in subsequent years.
This is likely to be a result of the development of protective antibodies following the first exposure. Repopulation of hares in a farm 3—4 weeks after an outbreak, to replace the hares that had died, resulted in death of the new hares in 1 to 2 weeks after introduction. The source of the infection, whether carrier hares that had survived or the contaminated environment, was not known.MOLECULAR EPIDEMIOLOGY
A French study on nucleotide sequencing of the partial capsid protein genes from EBHSV from different parts of the country between 1989 and 2003, and in comparison with a Greek strain, showed high level of conservation between EBHS viruses — the maximum nucleotide divergence was 11.7%(18). The study revealed two major groups and several genogroups, and showed that old strains of EBHSV persist and evolve slowly in the locality of their origin. Several different genogroups co-exist and there is apparently an efficient dissemination of new genogroups(18).
The emergence of EBHS is not understood. It has been postulated that low or apathogenic EBHSV strains might have occurred in Europe before the disease was detected. A phylogenetic study indicated that EBHSV originated in Sweden and spread to western, eastern and southern parts of Europe1-19). Increasing numbers of EBHSV strains are being identified by molecular investigations; 24 strains are presently known(19).
HOST FACTORS
Early studies in Sweden diagnosed EBHS in 275 out of 2818 European brown hares and mountain hares examined by histopathology between 1980 and 1989. The majority (70.5%) of the EBHS cases occurred in adults (hares with fully developed gonads), and the rest in subadults (hares whose gonads were not fully developed); no cases were found in lactating hares. According to the anamnesis, 88.2% of the hares were found dead, 5.9% were moribund and euthanized and 5.9% showed abnormal behaviour and died after a short clinical course.
It was later confirmed in several countries that EBHS mortality occurs only in adult and subadult hares. No cases are described in hares younger than about 50 days, either naturally exposed (during outbreaks) or experimentally infected, indicating that they are resistant. The mechanisms of the age-related resistance are not known. Males and females appear to be equally susceptible to EBHS.
Genetic analysis of hares to detect possible familial susceptibility to EBHSV identified 15 different matrilinear
hare haplotypes, but they were not correlated to occurrence of EBHSV or antibodies1-19).
TRANSMISSION
EBHSV is transmitted directly or indirectly. The virus is shed in faeces (probably transported from the liver, through bile into the intestine) and hares can be infected experimentally by the oral route. It is believed that the most important route of infection is the faecal-oral, and possibly the respiratory, but no specific comparative studies on susceptibility to infection by various routes and viral doses have been conducted. EBHSV is also considered to be transmitted via fomites.
PATHOGENESIS, PATHOLOGY
AND IMMUNITY
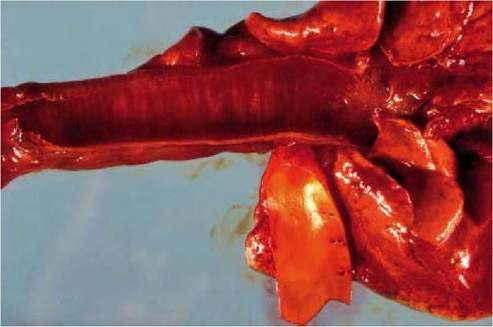
FIGURE 5.5 Lung, trachea and aorta of a hare with EBHS. Lung congestion and haemorrhages and congested tracheal mucosa. Froth in the opening of bronchi reflect lung oedema. The aorta has been opened to show yellow discoloration indicative of jaundice. Photo: Bengt Ekberg, SVA.
Little is known about the pathogenesis of EBHS. Most experimentally infected hares die 2—3 days post-infection (dpi); some hares die later, up to about 7dpi, and some hares survive. The liver is the main target organ, but the virus disseminates systemically. In terminal stages, hares develop disseminated intravascular coagulation and die with circulatory collapse, showing congestion in the lungs and other organs.
Characteristic gross lesions are fatty and friable liver, and signs of circulatory collapse, such as lung congestion and oedema, congestion of the mucosa of the trachea (Figure 5.5), and an enlarged and congested spleen.
Stomachs well filled with ingesta and catarrhal enteritis in the small intestine, with thick mucus in colon and rectum, are often observed. There may be haemorrhages in any organ, but frequently on the mucosa of the stomach and in the lungs(15). Signs of jaundice are observed in about one-third of the cases and are particularly frequent in subacute cases (Figure 5.5).Histologically, in most cases, the acute development of liver lesions reflects the rapid course of the disease. The predominant, consistent and characteristic lesion is periportal hepatocellular coagulation necrosis (Figure 5.6), which coincides with the localization of EBHSV in hepatocytes in those areas. To the knowledge of the author, no other diseases cause this particular zonal liver necrosis in hares. The reason for the preferential periportal distribution of lesions and virus is not known. While liver necrosis occurs in all hares that die of EBHS, its extent varies, from
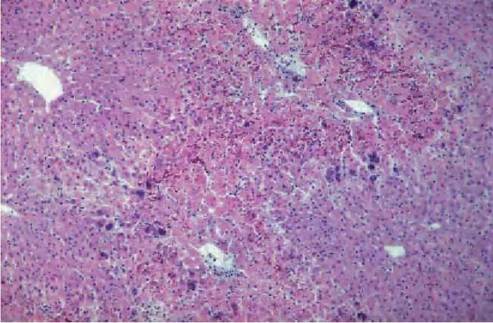
FIGURE 5.6 Histological section of liver of a hare with acute EBHS. Hepatocellular coagulation necrosis of periportal areas. Centrolobular (areas around the central veins) are spared and unaffected by the necrosis. Dystrophic calcification of hepatocytes is present. Inflammatory cell infiltrate is minimal. Photo: D. Gavier-Widen, SVA.
a small periportal area (Figure 5.6), to the whole lobule (massive necrosis). There is also lytic necrosis with a loss of hepatocytes, leaving a few heterophils in an empty reticulin framework. In the peracute form there is massive liver necrosis, haemorraghes and scant or no inflammatory infiltrate. Viral antigen is located in the nucleus and cytoplasm of hepatocytes. In subacute cases there is periportal hepatocellular necrosis, fatty degeneration, infiltration of
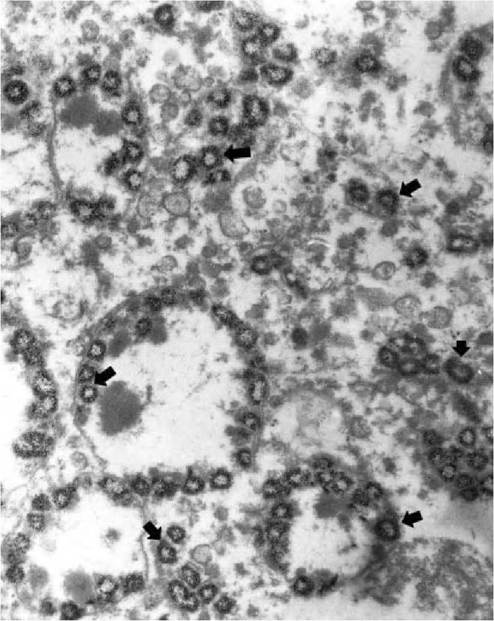
FIGURE 5.7 Electron micrograph of one hepatocyte liver of a hare with European brown hare syndrome. Mitochondria in the hepatocyte contain amorphous densities in the centre and large amounts of calcium as hollow spherules (arrows). Enlargement 9,000?. Photo: D. Gavier- Widen, SVA.
inflammatory cells and presence of virus in hepatocytes, which is more abundant in areas contiguous to the necrosis. Viral antigen is also present in the cytoplasm of macrophages (Kupffer cells); in cases with a longer clinical course of disease this is the predominant localization of virus. Acidophilic bodies, formed by condensation of degenerating hepatocytes, are often observed in sinu- soids(15). Granules of dystrophic calcification can be abundant in hepatocytes (Figure 5.6). They are formed by mitochondrial mineralization (Figure 5.7), and can be demonstrated with von Kossa staining in liver sections, even in autolysed cases. Chemical analysis of these livers reveals that the content of calcium is up to 20 times higher than that in normal hares(15). With a longer clinical course, there is more marked inflammatory infiltration, bile duct proliferation and hepatocytic regeneration, manifested by the presence of binucleate hepatoctes. A small proportion of the hares develop chronic hepatitis and harbour virus in the liver, although at lower levels than in acute cases, for a period of unknown length.
About one- third of the hares develop kidney lesions, with tubular cell degeneration, necrosis and in some cases mineralization. There is lymphoid cell depletion or reduction in lymphoid organs, such as the spleen and lymph nodes(15).
An antibody response to EBHSV develops early after infection, at about 5 dpi. Hares that survive apparently develop life-long immunity, but the course of antibody levels during long periods of time are not known. Newborn hares may have antibodies to EBHSV, indicating passive transfer of immunity. Young hares (younger than about 50 days) with no maternal immunity that are exposed to the virus will develop antibodies but not clinical disease.
CLINICAL SIGNS AND TREATMENT
Hares may die peracutely without signs of disease (Figure 5.8). In cases with a longer clinical course of disease the hares may show nervous signs, which are probably the result of hepatic encephalopathy. The hares frequently lose fear of people, run in circles, are apparently blind, show loss of balance, opisthotonus, adopt abnormal postures, and terminally cramp and lie in prostration(15). The hares die about 1—7 days after the first manifestations of clinical signs.
Experimentally, hares develop leucopenia by 24 hours post- infection, with values as low as 500 leucocytes per microlitre. Enzymes that indicate liver damage (aspartate aminotransferase and alanine aminotransferase) and bilirubin increase rapidly by day 4 post-infection, shortly before death. These changes are less marked in hares that survive the infection and reach normal values at day 10 post-infection(20). Analysis of urine shows proteinuria, casts and haemoglobinuria(15).
Treatment is normally not feasible, but antiserum given early in the incubation period may prevent death.
DIAGNOSIS
Gross lesions are not always observed. The histological features of the liver (periportal necrosis) are characteristic of EBHS but are difficult to identify in cases with autolysis. To detect virus, the liver is the organ of choice, as it

FIGURE 5.8 European brown hare syndrome outbreak; dead adult brown hares collected in one day from an estate in southern England in October 1991. Photo: J. Paul Duff. AHVLA. Disease of Wildlife Scheme.
has the highest titres, but the spleen often contains EBHSV The virus can be demonstrated by immunohistochemical or immunofluorescent staining of liver sections, and by haemagglutination (human type A or O red blood cells at pH 6.4 and at 4°C), negative staining electron microscopy of liver homogenate and western blot. Several antigendetection ELISA methods have been developed and used widely. Immunocapture reverse transcriptase PCR (RT- PCR), based on genomic amplification of the virus captured on a plate coated with anti-EBHSV antibodies has been reported as a rapid method of high sensitivity. Several PCR techniques have been developed for the detection of EBHSV nucleic acids in fresh or formalin-fixed, paraffin- embedded tissues.
Serological tests are usually ELISA methods. Antibody detection ELISA can be conducted on blood collected on blotting paper, but with lower sensitivity than on serum. Haemagglutination inhibition can also be used for serology. Antibodies to EBHSV and RHDV show a low level of cross-reaction.
MANAGEMENT, CONTROL AND REGULATIONS
Eradication of EBHS in free- living hares is not feasible. Most hare restocking areas in Europe have endemic EBHSV, and translocation or import of hares from these areas into free areas should be avoided. In hare farms the safeguards for the prevention of virus entry include quarantine, serologic testing of new hares before introduction to the farm, and fencing to avoid contact with wild hares, using processed feeds rather than field-collected grass. Indirect infection through contaminated hay, or clothes and shoes should be avoided. Autogenous killed virus vaccines can be prepared at the beginning of an episode of EBHS mortality in farms to vaccinate the still- healthy hares.
PUBLIC HEALTH CONCERN
EBHSV does not affect humans.
SIGNIFICANCE AND IMPLICATIONS IN ANIMAL HEALTH
EBHS can have a significant impact during initial outbreaks or in sporadic outbreaks, and local declines of hare populations have been attributed to EBHS. Some reports describe recovery of populations within a few years because of the high reproductive rate of hares and the immunity acquired by the population resulting in lower EBHS mortality. EBHS was the most common disease of mountain hares in regions of Finland(14) and over much of Europe it remains a frequent disease of the European brown hare. EBHS causes significant losses in hare farms.
More on the topic EUROPEAN BROWN HARE SYNDROME:
- 8 species (Family)-Specific wildlife DISEASES IN EUROPE
- 1 SOME WILDLIFE RELATED EMERGING DISEASES (WIREDS) IN EUROPE
- RABBIT HAEMORRHAGIC DISEASE
- Bibliography for viral infections
- 5 Appendices
- Pasteurella infections in wild mammals
- CHAPTER 34 Coxiella burnetii infection
- Index
- LATENT INFECTIONS (APPARENTLY HEALTHY CARRIERS)