Transposon Mutagenesis: Himarl vs. Tn5367 and Tn5370
The thickness and impermeability of the cell wall makes it difficult to introduce plasmid vectors into MAP. Therefore, the shuttle plasmid vectors described in Table 7.1 yield low transformation efficiencies.
To overcome this challenge recombinant shuttle phasmids were developed to introduce allelic exchange substrates and transposons into mycobacteria via the phage receptor to allow for efficient transfer of genetic material through the cell wall. These phasmids are also thermosensitive (e.g. phAE87) for the downstream recombination and selection processes that ultimately lead to gene inactivation and elimination of phage specific sequences (Chacon and Barletta, 2010).Two types of transposons have been extensively used to generate random mutant banks: (i) IS1096- and (ii) Himar1 -derived transposons (Fig. 7.1). Tn5367 (GenBank Accession KM232614) was the first transposon widely used in mycobacteria including MAP (Bardarov et al., 199 7; Shin et al., 2006; Rathnaiah et al., 2014). This transposon carries a kanamycin-resistant drug marker (aph), and the IS1096 transposase (tnpA) and resolvase (tnpR). This transposon has the transposase within the transposed element (Bardarov et al., 199 7) resulting in a relatively high transposition frequency (~1.0 ? 10-5), at least in Mycobacterium smegmatis (Cirillo et al., 1991). Tn5367 transposes by a cut and paste mechanism (McAdam et al., 2002). This element
Table 7.1. Previously reported genetic systems and strategies used in Mycobacterium avium subsp. paratuberculosis (MAP) mutagenesis and gene expression studies (1993-2008).
| Component | Description | Examples |
| Genetic systems Shuttle plasmids | Escherichia Coli-Mycobacterium replicative vectors used to introduce extrachromosomal copies of genes via transformation | pMV261 and pMV262 (Stoverefa/., 1991; Foley-Thomasefa/., 1995)pSMT1 (Rosseels etal., 2006) |
| Integrating plasmids (see Section | Integrase and L5 mycobacteriophage attP site | pMH94 (Leeefa/., 1991) pMV361 (Stover |
| 7.5 for use in MAP) | carrying vectors used to introduce DNA into the mycobacterial chromosome via transformation at the attB locus | ef al., 1991; Hatfult and Sarkis, 1993) pMV306 (Hahn ef al., 2005) |
| Phages and shuttle phasmids | Chimeras of a thermosensitive mycobacteriophage and an E. coll cosmid used to introduce DNA into the MAP chromosome via transduction | phAE87 vector and phAE94 carrying Tn5367 (Bardarovefa/., 1997; Harrisefa/., 1999) |
| Reporter genes | Firefly and bacterial luciferase genes introduced into MAP via shuttle phasmids or plasmids Green fluorescent protein gene introduced into MAP via shuttle plasmids β-galactosidase promoterless truncated gene introduced via shuttle plasmid. Transcriptional/ translational fusions used to identify MAP promoter consensus sequences and determine the strength of promoters and ribosomal binding sites | phAE39 (Foley-Thomasefa/., 1995) phAE85 (Sasahara ef al., 2004) pSMT1 (Rosseels ef al., 2006) pYUB180 (Williamsefa/., 1999) pWES4 (Parkerand Bermudez, 1997; Harris ef al., 2002; Park ef al., 2008) pYUB76 (Barlettaefa/., 1992; Bannantine ef a/., 1997) |
Continued
G. Rathnaiah eta∕.
Table 7.1. Continued
| Component | Description | Examples | |
| Genetic manipulation | Transposon mutagenesis | Transposon Tn5367 introduced via the | /WAPstrain K-10 (Harrisefa/., 1999; Harris |
| strategies | thermosensitive phasmid phAE87 or a shuttle plasmid used to generate and screen comprehensive mutants libraries | and Barletta, 2001) MAP strains 989 and TMC1613 (Cavaignac etal., 2000) MAP strain ATCC® 19,698 (Shin ef al., 2006; Alonso-Hearn etal., 2008) | |
| Gene replacement | Substrates for homologous recombination introduced via shuttle thermosensitive phasmids (phAE87) and plasmids used to inactivate specific MAP genes | pknG, relA, Isr2 MAP K-10 mutants (Park ef al., 2008) IipN MAP K-10 mutant (Wu ef al., 2007) | |
| Complementation | Wild-type gene introduced into mutant strains via a multi-ńîđó shuttle plasmid carrying apramycin- resistant marker | pMAV261 -∕WAP~3464 and pMAV261- ∕WAP-3464, ∕WAP-3465 and ∕WAP~3466 (Alonso-Hearn ef al., 2008) |
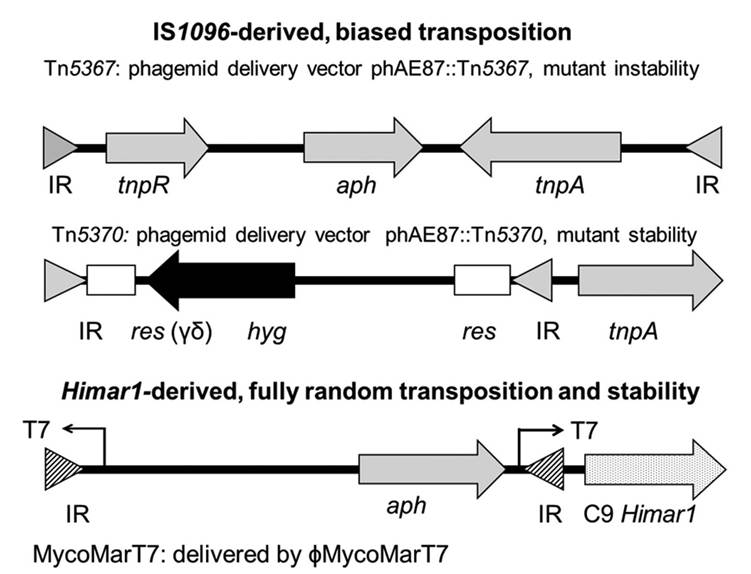
Fig.
7.1. Structure of mycobacterial transposons utilized to generate random mutant libraries. Inverted repeat (IR) for Tn5367 or Tn5370 (filled triangle), or Himarl-derived transposon (striped triangle); tnpR, IS1096 resolvase; aph, kanamycin-resistant gene; tnpA, IS1096 transposase; res, resolution site for transposon γ-δ; hyg, hygromycin resistant gene; and C9 Himarl, Himarl transposase. (Figure reproduced from Frontiers in Veterinary Science; Rathnaiah et al., 2017.)may also insert into another gene creating a new mutation and either a wild-type or mutated gene, depending on precise or imprecise excisions, in the location of the original insertion. Thus, the corresponding MAP mutants may not be stable enough to conduct long-term in vivo studies where bacilli multiply to large numbers. An improved IS1096-derived transposon, Tn5370 (GenBank Accession KM232615), was constructed by removing open reading frames (ORFs) dispensable for transposition (e.g. tnpR) (McAdam et al., 2002). Moreover, the trans- posase was engineered into the phasmid outside of the transposable element. Thus, Tn5370, as opposed to Tn5367, generates stable mutants upon transposition. This transposon has an additional advantage of possessing a hygromycin (hyg) marker outflanked by the resolution sites (res) of the γ-δ resolvase (Wiater and Grindley, 1988). This construction allows for the excision of the drug marker and the generation of unmarked transposon insertion mutants. However, none of these transposons inserts in a fully random manner in mycobacteria (McAdam et al., 2002; Shin et al., 2006).
Himar1-derived transposons have also been used to generate random mutant libraries in Mycobacterium tuberculosis and more recently in MAP (Sassetti et al., 2001; Scandurra et al., 2009; Rathnaiah etal., 2014; Wang etal., 2014). The Himar1 transposon (MycoMarT7; GenBank Accession AF411123) was engineered into the shuttle phasmid phAE87 (Table 7.1). This vector carries the highly active C9 Himar1 transposase outside the inverted repeats, thus yielding stable mutants.
In addition, this vector carries the aph drug marker and a T7 promoter that reads outward to facilitate identification of insertion sites by in vitro transcription and complementary DNA (cDNA) synthesis (Sassetti etal., 2001). The major advantage of the Himarl derivative is the almost fully random recognition sequence (5'-TA-3') vs the recognition sequence of IS 1096-derived transposons (5'-NNPy(A∕T)A(A∕T)NN-3') showed experimentally to have a significant transposition bias in both M. tuberculosis and MAP (Shin et al., 2006; Rathnaiah etal., 2014).Most of the MAP transposon mutant libraries were made with Tn5367 (Harris et al., 1999; Cavaignac et al., 2000; Shin et al., 2006; Rathnaiah et al., 2014) and fewer libraries have been constructed using the Himar1-derived transposon MycoMarT7 (Scandurra et al., 2009; Wang et al., 2014; Rathnaiah et al., 2016). A recent study performed a comparative analysis of MycoMarT7 and Tn5367 transposon insertion sites in MAP (Rathnaiah et al., 2016). They correlated the analysis of transposon recognition sites with actual insertions obtained in mutagenesis experiments to determine potential transposition biases. The analysis indicated that the distribution of IS 1096 insertion sites within ORFs is sparser, with 710 out of a total 4350 (16%) potential coding sequences containing no recognition sites. The corresponding mutants in those ORFs will not be observed in the mutant libraries irrespective of whether the gene is essential or non-essential in the conditions studied. In contrast, there are only 37 coding sequences not targeted by the Himar1 transposase. Thus, Tn5367 or Tn5370 may not be used for a comprehensive analysis of virulence determinants and gene essentiality in MAP. Most importantly this study also revealed transposition biases that are independent of the transposon site recognition sequences. Both transposons have loci-dependent biases, with Tn5367 being even more skewed. Most notably, insertions into categories of loci with fewer recognition sites seem underrepresented compared with the expected numbers of total sites.
Moreover, Tn5367 has an increased predilection for insertion within intergenic sites. These biases may explain the relatively high number of mutants with interesting phenotypes previously reported with insertions within intergenic regions (Rathnaiah et al., 2014). These loci-dependent biases may lead to an underestimation of the number of independent mutants required to generate a comprehensive mutant library. This indicates that a collection of at least 400,000 mutants would be required for a representative MAP Himar1 library. This number exceeds the 100,000 estimated for M. tuberculosis (Bardarov et al., 1997). For Tn5367, considering its predilection for intergenic regions and loci biases, a comprehensive library may need a collection of ~250,000 mutants rather than 13,500 as previously estimated based on random transposition (Harris and Barletta, 2001).Himar1 transposons have been used in MAP to isolate 111 mutants attenuated in bovine monocyte-derived macrophages from a random screen of 2290 (Scandurra etal., 2009). Though a sensible genetic approach was used, the screening method is labour intensive to screen a representative mariner transposon library. More recently, a larger study was performed that collected 90,000 Himar1 mutants but identified insertions in approximately 12,000 out of 52,000 (~25% saturation density) recognition sites (Wang et al., 2014). The authors attributed this low number of recognition sites represented to bottle-necking effects during DNA sample preparation or the sequencing stage. Loci-dependent transposition biases may also contribute to this effect. None the less, this Himar1 library was used to screen for mutants capable of conditional growth without mycobactin (Wang et al., 2016). Based on this property, they identified a potential mycobactin-independent iron uptake system on the MAP-specific genomic island LSPP15. Step-by-step procedures to generate a MAP transposon mutant library are detailed elsewhere (Bannantine et al., 2019).
7.5
More on the topic Transposon Mutagenesis: Himarl vs. Tn5367 and Tn5370:
- Concluding Remarks
- Cleft Lip/Palate
- GENOMIC CONSIDERATIONS FOR THE PATHOLOGIST
- Introduction
- IN VITRO EFFECTS OF PR MUTATIONS ON CELL KILLING
- AN OVERVIEW OF CLASSIFICATION AND STRUCTURE OF MAMMALIAN DEFENSINS:
- References
- Boon Andrew. The Ethics and Conduct of Lawyers in England and Wales. Hart Publishing,1999. — 808 p., 1999
- Griffiths-Baker Janine. Serving Two Masters: Conflicts of Interest in the Modern Law Firm. Hart Publishing,2002. — 227 p., 2002
- Grisso T.. Evaluating Competencies: Forensic Assessments and Instruments. 2nd edition. — Springer,2002. — 564 p., 2002