Apparent competition
Apparent competition is almost always present when one or more consumers exploit two or more resources. It is the interaction between the resources via those shared consumers (prey via shared predators).
Apparent competition has already been mentioned in a variety of contexts in both this and preceding chapters. Many of the same models used to understand competition can be used to understand apparent competition. Apparent competition shares some of the definitional problems associated with interspecific competition. Robert Holt's (1977) article, which introduced the term ‘apparent competition, noted that commonly used measures of the effect of one resource on another could be positive rather than negative, and most authors have subsequently referred to such cases as ‘apparent mutualism' (e.g., Abrams et al. 1998; Holt and Bonsall 2017). Mutually positive effects seem particularly likely when consumer-resource systems undergo self-generated cycles (Abrams et al. 1998). This work also noted the possibility of (+,-) effects between resources that share a consumer. If ‘competition’ is used for all interactions involving shared resources, consistency argues for applying ‘apparent competition’ to all interactions via shared consumers (including parasites and diseases), regardless of the attendant set of changes in population size following neutral perturbations to each species. This is a question for the ecological community to decide.Another diverse set of authors has argued in favour of dropping the ‘apparent’ and using the general term ‘competition’ to apply to both interactions via shared resources and shared natural enemies. Nicholson (1937) and MacArthur (1972) had mentioned shared enemies as a type of competition, and, more recently, Kuang and Chesson (2008) argued that these two indirect interactions were fundamentally the same in terms of coexistence.
Kuang and Chesson’s (2008) assumption of unrepresented resource species that would sustain the predators in the absence of the ones that are explicit in the model means that they treated the two trophic levels in a fundamentally different manner. As shown later in this section, there are some important differences between competition and apparent competition in a simple 2-consumer- 2-resource case. While it is true that both competition and apparent competition can cause exclusion of one or more species on the focal trophic level, there are differences due to the directionality of energy flow, which goes upward from lower to higher trophic levels in food webs. The resource level can exist without their consumers, but the predators on those consumers cannot do so. This fundamental fact would argue for a distinction between the two processes, but there are several other important differences that are noted below. The effects of assuming type II rather than type I (linear) functional responses differ qualitatively depending on which trophic level is affected. The same is true of the role of increasing direct density dependence in the transmitting entity/entities. In addition, the fact that consumer behaviour changes in different ways in response to changes in the abundances of resources and predators is another reason for distinguishing between the two interactions.As was noted above, theory regarding apparent competition has never displayed any tendency to leave out the dynamics of the predator of the indirectly interacting prey. Neither Lotka nor Volterra used what we now call apparent competition as an example of the phenomenon they were modelling. The importance of such factors as predator self-regulation, nonlinear predator functional responses, and unstable population dynamics have been noted repeatedly (Holt 1977,1984; Holt and Kotler 1987; Abrams 1987d, Abrams et al., 1998), and were reviewed recently (Holt and Bonsall 2017). It has long been recognized that a shared predator can produce mutually positive effects between two prey species (Holt 1977; Holt and Lawton 1994; Holt and Bonsall 2017; Abrams 1987d; Abrams and Matsuda 1996).
These articles show that type II or other nonlinear functional responses are an important mechanism by which such ‘apparent mutualism’ comes about in models of shared predation. This mutually positive outcome does not require consumer-resource cycles; it occurs whenever a type II predator also has some additional limiting factor other than its prey (e.g., good hunting sites). Nevertheless, population cycles often contribute to mutually positive effects via a shared predator.A type II response in a 1-predator-1-prey system produces larger amplitude cycles when the predator can persist on a low abundance of prey, and when there is a relatively large handling time. The addition of a second prey with a similar handling time usually increases the amplitude of the cycles, and higher amplitude cycles, whatever their cause, tend to increase mean prey density. Thus, (+,+) interactions are relatively common when the interaction sign is determined by the difference in mean prey density between systems with one vs two prey species. However (+,-) effects may also occur when prey differ significantly in handling time or some other properties (Abrams et al. 1998).
Although theory regarding apparent competition has always incorporated explicit dynamics for the effect-transmitting species, it has suffered from some of the same deficiencies as theory on resource competition. Systems with more than one predator or more than two prey are seldom examined, and behavioural adaptations of the species involved are also rarely considered. The remainder of this section will provide a brief exploration of a few of the implications of these omissions.
A system with two consumers that are limited solely by two dynamically independent resources has been a staple of competition theory. As a result, one might think that it should have played a role in theory regarding apparent competition as well, since it involves both resources sharing both predators. However, this food web is virtually absent from the literature on apparent competition.
This is because of a somewhat inconvenient truth. If the indirect interaction between the two resources is measured by the effect of a small change in the neutral parameter (ri in eqs (5.5) in Box 5.1), one finds that there is no effect of either resource species on the other resource’s equilibrium abundance at a stable equilibrium. (This is true provided the consumers have no limiting factors other than the two resources.) The equilibrium resource abundances are determined entirely by the consumer’s parameters in this case (see Box 5.1, which deals with logistic resources and type II consumers). However, a sufficient increase in one of the resource intrinsic growth rates, ri, will cause extinction of the predator which is more dependent upon the other resource. Similarly, a sufficient decrease in one ri will cause extinction of the consumer that has a greater relative consumption rate of that resource. Extinction of one of the two consumer species leaves the system with a single consumer species, so it will display apparent competition with mutually negative effects under the normal circumstances for such a model (for example, all cases with a stable equilibrium in which both resources have positive densities). A sufficient reduction in ri will cause extinction of resource i itself, leaving a simple 1-consumer-1-resource system, usually with a higher abundance of the remaining resource. Note that the independence of the equilibrium populations of the two resource populations (over some range of ri) only depends on having a stable equilibrium and having consumer per capita growth rates that are solely dependent on the two resource abundances. Thus, adding consumer switching behaviour or changing to any other set of functional responses that are solely determined by resource abundances would not affect the lack of a between-resource interaction.Box 5.1 Interactions in a potentially unstable 2-consumer-2-resource system
'The model of two consumers and two resources used here is:
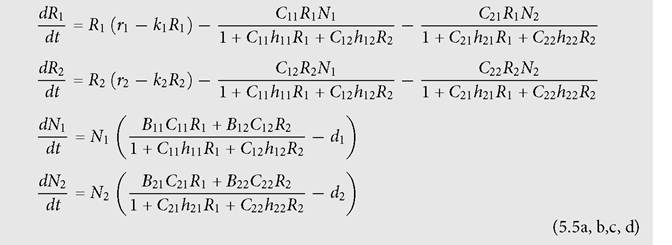
In the simplified example, the two resources have equal growth parameters, all of the Bij are identical to each other, as are the hij; in addition, d1 = d2 in the system before any perturbation.
The consumers are assumed to have mirror image consumption rates, with C11 = C22 = C, and C12 = C21 = 11 - C, where 1/2 < C prevents apparent competition, only depends on the absence of terms involving consumer abundance in the expressions for consumer per capita growth rate. If consumer densities directly affect their own functional responses, this independence will no longer hold.The consequences of mortality (reduced r) of one resource species for the population size of the other are more complicated if the system exhibits sustained cycles before any perturbation. In this case, we can again get some general insights by examining a symmetric system having ‘mirror image’ competitors using the same two resources that initially have identical growth parameters, but differ by having opposite susceptibilities to being consumed by the two consumers. The interaction between the resources is again measured by a change in their mean densities following a change in the neutral parameter ri of each on the other. Mortality applied to either resource (lowering ri) reduces the amplitude of the population cycles.
Lower amplitude cycles in this system are detrimental to both resources because the mean consumption rate increases when the consumers spend less time handling resources. As a result, both resources decline in mean abundance in response to a lower r of a single resource for many parameter sets. This phenomenon is influenced by the degree of difference between the two consumers in their relative consumption rates of the two resources. It can also be affected by the presence of switching predation. Hydra effects in the resource (increases in the mean density of a resource with a decreased r) occur in some cases, and the between-resource interaction has a complicated, nonlinear form. As noted above, a sufficient decrease in r (e.g., increase in the mortality) in one resource will cause the extinction of the consumer that is more specialized on that resource.
Once that happens, the interaction (now in a 2-resource-1-consumer system) often reverts to apparent competition. However, if the cycles are still relatively large in amplitude at this point, apparent mutualism is still possible (Abrams et al. 1998). Further increases in the mortality of one resource will eventually cause its extinction, leaving a 1-consumer-1-resource system.Figure 5.7 illustrates the indirect interaction between the two resources in a potentially unstable version of the 4-species web described in Box 5.1. The figure illustrates the mean abundances of the two resources as a function of the intrinsic growth rate of one of them (r1). Because of the symmetrical structure of the community, the effects of changing r2 on the two resource abundances are identical (with resource identities reversed). Figure 5.7A has a moderate degree of resource partitioning between consumers (C = 0.7), while panel B of the figure assumes a higher level of resource partitioning (C = 0.8). In both panels, the baseline case assumes equal intrinsic growth rates of the two resources (r1 = r2 =1). The main message from the figure is that, except for values of r1 that are low enough to cause extinction of consumer species 1, the interaction of the two resources via the consumers often cannot be described as mutually negative. In addition, the presence of cycling implies that there is normally some effect of a change in the resource per capita growth rate on the mean abundances ofboth resources. (Recall that models of the form of eqs (5.5) with a stable equilibrium exhibit no effect of a changed neutral parameter of one resource species on the equilibrium abundance of the other.) An increase in the r-value for resource 1 increases the mean abundances ofboth resources over the part of the parameter range illustrated in Figure 5.7A in which all four species are present. The expected lack of effect on the equilibrium R2 does not occur when r1 is increased because the higher amplitude of consumer-resource cycles increases the mean level of consumer satiation, and therefore decreases the consumers’ net effect on both resources. The symmetry of the original system guarantees that a reduction in r2 would have the same relative effects as reducing r1 by the same amount; resource 2 would increase in population size and resource 1 would increase to a smaller extent. Although abundances for r1 > 1.2 are not shown, both resources continue to increase with a larger r1 throughout the range of r1 values that allow predator 2 to exist (it becomes extinct for r1 slightly above 2.20).
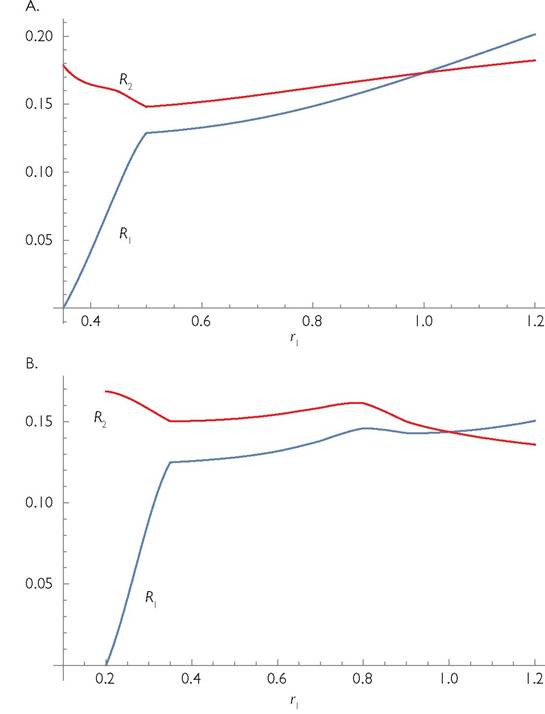
Fig. 5.7 Apparent competition in 2-consumer-2-resource systems with type II consumer functional responses. The model (eqs 5.5) is given in Box 5.1. Each panel shows the effect of changing the maximum per capita growth rate of resource 1, r1. The two resources have identical r’s, and therefore identical equilibrium and mean abundances when r1 = 1. The parameters in the system illustrated here are: r2 = 1, k1 = k2 = 1; hj = 2; Bj = 1; d1 = d2 = 0.1. Panel A assumes that Cii = 0.7 and Cj = 0.3; panel B assumes greater resource partitioning by the consumers; Cii = 0.8 and Cij = 0.2. In panel A, the abrupt change at r1 values slightly below 0.5 corresponds to loss of consumer 1 at that point. Resource 1 is extinct if r1 ≤ 0.35. Both resource species increase in mean abundance as r1 is increased above 0.5. In panel B, resource 1 is extinct for r1 < 0.2, and consumer 1 is extinct for r1 values below approximately 0.35. The system exhibits cycles for the entire range of parameters shown.
The greater level of resource partitioning assumed in panel B of Figure 5.7 produces a somewhat different range of interspecific effects between the resources, but there is again some interaction over the entire range of r1 shown (and for larger values that are not shown in the figure). The apparent competition at low r1 reflects the fact that consumer 1 is absent. Following the entry of consumer 1 to the system, further increases in r1 produce relatively weak positive effects on both species. In Holt's (e.g., Holt and Bonsall 2017) terminology this would be categorized as apparent mutualism. Still higher values of r1 (> 0.8) are characterized by mutually negative changes in resource abundances following a small increase in r1. The negative effect of r1 on the abundance of resource 1 constitutes a hydra effect, so the interaction with the other resource could be considered positive or negative depending on whether the perturbation to resource 1 is regarded as positive (a higher per capita growth rate) or negative (a lower mean population size). For r1 > 0.9, greater r1 again increases the mean R1 and decreases the mean R2, so the interaction is again apparent competition. Although it is not shown in the figure, the effect of greater r1 becomes positive for both resource abundances for r1 slightly above 1.4.
The qualitative pattern of change in the two resource densities with altered per capita growth (larger or smaller r) of one of them, shown in Figure 5.7, differs for other levels of resource partitioning between the consumers (different values of C), which are not illustrated. If C = 0.5, the two consumers are identical generalists, so they are unlikely to coexist. This reduces the system to the 1-consumer version of eq. (5.4), which was discussed in Abrams et al. (1998). The 1-consumer version does have some parameters producing mutually positive effects, although they are less prevalent than in the 2-consumer case with cyclical dynamics illustrated here. Relatively low levels of resource partitioning (e.g., C = 0.6) produce mutualistic interactions between the two prey, but a relatively modest reduction in r1 leads to the exclusion of predator 1 and stabilization of the system, so further reductions in r1 will increase the equilibrium R2. Very large values of C (close to unity) imply near-specialized predators, so there is very little indirect interaction between them.
There is much to be learned about interactions between two or more prey that share two or more predators. However, the common assumption of mutually negative effects is clearly far from universal. One topic that has received relatively little attention is the interaction between biotic resources in systems where saturating functional responses may produce cycles and the consumers also display adaptive switching. This is of interest because switching can increase the range of parameters that allow persistence of all of the species; i.e., it greatly reduces resource exclusion caused by apparent competition. The effects of switching can also be like those of adaptive evolution of the consumer's resource-capture abilities, when there is a trade-off between greater consumption abilities of different resources (as in Abrams 2006b). Preliminary numerical exploration of eqs (5.5) with the addition of the switching function (eq. 5.4) suggest that in cycling systems the positive effects of an increase in the r value of one resource on the abundance of a second resource is usually significantly larger than in a comparable model without switching. The switching model also has positive effects over a larger range of parameter values. This maybe in part because of the increased synchrony in the dynamics of the two species on a single level, which tends to increase the amplitude of population cycles. Further exploration of this case, and other models of adaptive predation, should help clarify the role of adaptive processes in shaping apparent competition between two resources sharing two consumers.
Cases of shared predation (apparent competition) with both more than two prey and two or more predator species are also understudied. Holt's original (1977) article considered an arbitrary number of prey with a single predator, but did not examine cycling systems. Some cases with two-or-more predators and three-or- more resources are looked at to address specific questions in Schoener (1993), Kamran-Disfani and Golubski (2013), and Rossberg (2013).
In ending this chapter, it is interesting to look back at the question of whether apparent competition should just be considered as another type of competition. As noted previously, Kuang and Chesson (2008) and Sommers and Chesson (2019) have revived an old argument for considering both apparent competition and resource competition as similar feedback processes. It is certainly true that both predators and resources can limit the ability of a species on an intermediate trophic level to exist, but they have many differences in their mechanisms and evolutionary consequences (e.g. Abrams 2000a, Abrams and Chen 2002a; see Chapter 11). The lack of any interaction between resources when stable systems have equal numbers of consumer and resource species (and when there is no direct effect of consumer abundance on consumer per capita growth rate) represents another important difference between competition and apparent competition. All of this argues for the traditional practice of regarding resource-mediated and consumer-mediated indirect effects as different interactions, despite some obvious analogies in their mechanisms.
More on the topic Apparent competition:
- Contents
- Conclusions
- Consequences of nonlinear functional responses
- Reasons for including resource dynamics
- Resource extinction and quasi-extinction in MacArthur’s model
- Abrams Peter A.. Competition Theory in Ecology. Oxford University Press,2022. — 336 p., 2022
- Theory regarding the strength of competition
- Introduction
- Arguments against resource-based definitions and models
- Evolution with both exploitative and interference competition