Space and the global shape of intraspecific competition
Interspecific competition is always closely related to intraspecific competition, so it is logical to begin with a treatment of how spatial patch structure affects the nature of intraspecific competition.
In general, quantification of interactions defined at the level of a metapopulation (or metacommunity), requires changing a neutral parameter of a focal consumer species across all patches, and then measuring resulting changes in total equilibrium or mean population sizes. One of the most important descriptions of intraspecific competition is the shape of this relationship. If the patches differ from one another (which is normally the case in natural systems), a neutral parameter perturbation of the same magnitude across all patches will likely have quantitatively different effects on consumer abundances in the different patches. Because the per capita competitive effects in most patches change with consumer and resource abundances, this implies that the magnitude of intraspecific effects at the metacommunity level will change as well.The simplest case to explore is one in which migration is both random and very rare. The within-patch abundances may then be assumed to reach approximately the abundance they would have reached if the patch had been completely isolated.
I consider a multi-patch system having z patches, each obeying the 1-predator-1- prey MacArthur model, with both species potentially present in all patches. This is described by the following equations, where the subscripts denote patch of residence rather than species identity:
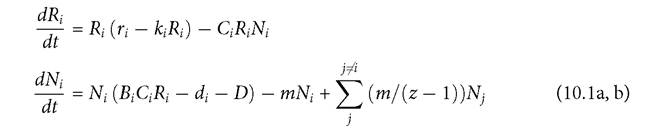
The parameter definitions are identical to those in the equivalent model in previous chapters. All consumers have equal per capita movement rates (m) out of a patch, and all individuals are assumed to survive the dispersal process.
Each disperser has an equal probability of arriving at one of the other patches. D is the perturbation to the mortality rate.Assuming that the movement rate m is small enough that it does not affect consumer abundance appreciably, the following approximation for the abundance of the consumer in patch i can be applied:


Note that, as the mortality perturbation, D, increases, consumer extinctions occur in the order given by the value of the right-hand side of inequality (10.3); this means that the patches i having the lowest consumer per capita growth rates when the resource is at its carrying capacity will be the first to drop out of the summation in eq. (10.4). These are patches that are characterized by low values of B, C, and/or r as well as those with high values of ki and/or di. High k, low C, and/or low B within a particular patch are all associated with a large change in Ni per unit change in D. Therefore, the consumer extinctions that occur at relatively low mortality perturbations are likely to reduce the total rate of decline of the total N with D by the largest amount. For a simple example, consider two patches that differ only in their values of C, with patch 2 having the smaller C, equal to 1/3 that of patch 1. The smaller capture rate means that the predator goes extinct in an isolated patch 2 at a lower imposed mortality than in patch 1. Patch 2 has a larger rate of decline of N with D than does patch 1. The result of extinction of N2 is that there is an abrupt decrease in the magnitude of the negative slope of N1 + N2 vs D at this point.
The formulas and the 2-patch example just discussed made the simplifying assumption that migration between patches was close to zero. This approximates the case of a very low rate of random movement between patches.
In the limiting case of very rapid (and cost-free) movement, the patches merge into a single one having the mean of the two patch-specific parameter values. The effects of most intermediate movement rates must be determined numerically. Figure 10.1 illustrates these effects for two cases with relatively low movement rates, assuming patches are identical except for patch 2 having a C value 1/3 that of patch 1. The blue dots represent a movement rate that is 33.3 times larger than the (otherwise equivalent) system represented by the red dots. Both cases are characterized by a positive second derivative of the relationship between Ntotaι and D. This relationship is linear for the MacArthur model in a homogeneous environment, or in a metacommunity model consisting of identical patches with MacArthur-type dynamics in each.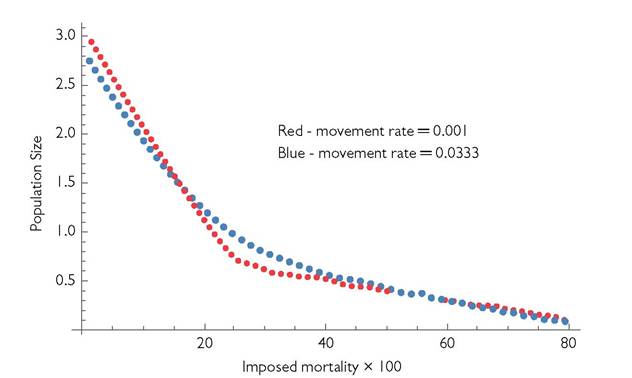
Fig. 10.1 Population size as a function of general mortality applied to the consumers in a 2-patch metacommunity with a single resource and a single consumer species. The parameters are: B1 = B2 = 1; C1 = 1; C2 = 1/3; r1 = r2 = 1; k1 = 1, k2 = 1; and d1 = d2 = 0.1.
If the above approach is expanded to a system having patches that differ in their values of two or more parameters, the same phenomenon of local consumer extinction occurs, and it causes a similar concave shape of Ntotat and D. This phenomenon is similar to what occurs with multi-resource systems in a single patch (see Abrams (2009b) and Chapter 5). If the resources in the individual patches have abiotic growth, the N vs D relationship is concave for a single isolated patch, and the concavity is accentuated by having local consumer extinction with increasing D. If biotic resources have theta-logistic growth with exponents other than 1, the individual segments of the relationship are nonlinear, so the entire relationship becomes more complex in its shape.
Random movement is unlikely in any species that is capable of self-directed travel. Chapter 3 argued that adaptive foraging was prevalent in natural communities, and this is often associated with directed movement. If resources are distinguished both by their physical identity and where they occur, then transfers of both consumers and resources between patches have key roles in determining the strength of indirect interactions among different consumers and among different resources. The next two sections consider the major distinguishing features of such movements; whether they are independent of consumer and resource abundances (‘random’) or depend on them in a manner that is adaptive for the moving entity.
10.4
More on the topic Space and the global shape of intraspecific competition:
- Intraspecific competition
- Understanding intraspecific and apparent competition
- Space and Scale between the Local, Regional and Global
- Though the rise of religious violence has been a global phenomenon in the modern period, perhaps nowhere is the arena of competition among contesting religious and secular politics greater than in South Asia.
- New Shape to the Old Demon
- Global Fiscal Policies Implemented Within the Scope of Fight Against COVID-19, a Global Public Bad
- Typical and unique shape of the state.
- CONCEPT 18.2 Global patterns of species diversity and composition are influenced by geographic area and isolation, evolutionary history, and global climate.
- Epilogue: The New Shape of the Industrial World The West can rise again.
- CHAPTER TWO The Shape-Shifting, Never-Changing World of Fraud
- Visual Space
- Our Space and Health
- Our Space and Education