Random movement and coexistence
This section assumes random movement of consumer individuals. As above, ‘random’ movement implies per capita movement rates that are independent of consumer and resource abundances and conditions in both the patch of origin and in all potentially accessible destination patches.
All resource items are identical, and there is strictly exploitative competition for this single resource type within each of the two patches. The simple models used here again do not allow alternative equilibria within a patch, unless an interference component to competition is added. However, having different physical conditions in the two patches often allows two consumers to persist in the system as a whole.The question is how greater movement of consumers and/or resources changes competition between consumers; this requires a measure of competition. Here I measure competitive strength using the change in the ‘existence bandwidth’ that is produced by the competitor. This is a simple extension of Armstrong’s (1976) ‘coexistence bandwidth’ (see Chapter 6) to purely intraspecific competition. The existence bandwidth is the range of density- and location-independent mortality rates (or the range of another neutral parameter) that allows a consumer to exist in the absence of the competitor. ‘Coexistence bandwidth’ is the corresponding measure in the presence of the competitor. The strength of competition can be measured by comparing the coexistence bandwidth to the existence bandwidth. This provides a measure that can be used to compare systems with different nonlinear competitive effects. If there is no competitor, the maximum increase in the death rate of one consumer (say the consumer in patch 1) is simply the maximum per capita growth rate from near-zero abundance in the patch where this consumer has its highest per capita growth rate; i.e., b1c1r1∕k1-d1 in the MacArthur model.
Competition is measured by the proportion by which the competitor’s presence at the joint equilibrium (or cyclic attractor) reduces this range.The following two cases are the simplest ones; i.e., those in which either consumer^) or resource(s) is/are unable to move.
10.4.1 Competition when only one trophic level moves
The first case considered here assumes density-independent resource movement at a fixed per capita rate, with each of the consumer species confined to a different patch. With no resource movement, the two consumers coexist globally. However, resource movement makes coexistence more difficult by making the food supply of the consumer that occupies patch 1 dependent upon consumption by the second consumer species in patch 2. The larger the per capita movement rate of the resource between patches, the greater dependence of the resource in patch 1 on consumption occurring in patch 2. A sufficiently high movement rate means that the resource population is nearly equal across patches, so that even small differences in the zero-growth requirements of the two consumers will result in exclusion of the less efficient one. The ability to coexist can be quantified by examining how the existence bandwidth, defined above, is changed by the presence of the competitor. Again, the MacArthur model provides a useful case for illustrating some numerical results. The question is: how rapid must resource movement be to make coexistence of patch-restricted competitors unlikely? The example discussed here assumes equivalent parameters for the consumers in the two patches, with a per capita resource movement rate of m between the two patches.
Figure 10.2 plots the proportional increase in consumer death rate required to cause extinction as a function of resource movement rate. The two lines illustrate this relationship for two different baseline consumer mortality rates. The red line corresponds to a baseline mortality of 0.25, and the blue line corresponds to a mortality of 0.10.
The maximum per capita resource movement rate shown on the figure is four, which is four times larger than the maximum per capita reproductive rate of the resource. At this point, coexistence is possible until the mortality rate disadvantage is approximately 1.3-fold in both cases illustrated; smaller disadvantages allow coexistence. Spatial separation can be effective at generating coexistence even with relatively rapid resource movement.The second case considered assumes that the resource individuals/units do not move between patches. However, the consumers have random movement between patches and have one or more growth parameters that differ between patches, therefore producing different competitive abilities in the two patches. I assume that a very low level of movement from outside this system ensures that both consumers and
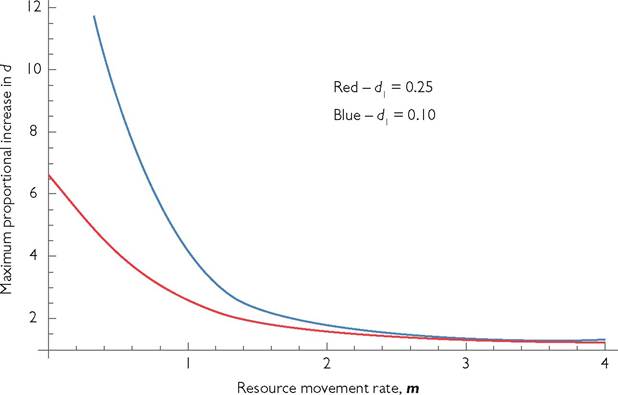
Fig. 10.2 Maximum proportional increase in death rate allowing existence for spatially separated competitors with resource movement at a rate m. Greater movement implies greater competition, reflected in a lower maximal increase in mortality. The common parameters are: Bi = B2 = 1/4; Ci = C2 = 2/3; r1 = r2 = 1; and k1 = k2 = 1/10. The blue line represents a system with d1 = d2 = 1/10; the red line is a system with d1 = d2 = 1/4.
the resource reach both patches. If there is near-zero movement between patches by the consumers, system-wide coexistence at a stable equilibrium only occurs if each consumer is the superior competitor in a different patch. Low rates of random movement of the consumers allow unequal coexistence in the two patches. High rates of random movement can result in exclusion of the less abundant or slower reproducing consumer.
Abrams and Wilson (2004) and Namba and Hashimoto (2004) carried out similar analyses of competition in this system.
Both articles showed that robust coexistence was possible in a 2-patch model with random consumer dispersal despite each consumer having identical equilibrium resource densities (R*) in each patch in isolation. Coexistence was also possible even when the same consumer was the superior competitor in each patch in isolation (i.e., one species had a lower R* in both patches). This consumer would normally be expected to exclude the other. However, coexistence is possible when the consumer with the exploitation disadvantage (higher R* values) has a lower movement rate or a faster demographic response to changed resource abundance (e.g., proportionally greater values of both resource conversion efficiency and per capita death rate). Either of these differences results in a greater concentration of the ‘inferior’ (higher R*) competitor in the patch with the greater resource density. This means that the consumer that is an inferior exploiter is also an inferior (slower) disperser.Another mechanism is observed when the system is continually perturbed away from an equilibrium that would otherwise be stable. If the system experiences randomly occurring consumer mortality events, then the slower disperser must usually be a better competitor (lower R*) in order for both consumers to persist in the system while competing for the same resource within each patch. This is the well-known competition-colonization trade-off, which requires that the patches experience perturbations that keep them from remaining long at near-equilibrium conditions. It does not require directed movement, but the presence of some directed movement is consistent with the mechanism. Rapid resource regeneration following a consumer mortality event usually makes the mechanism more effective in promoting coexistence.
While two species can coexist via the differential movement-rate mechanisms described above, it is unlikely that large numbers of species do so. Most cases of coexistence of multiple consumer species in a metacommunity that contains only one or a small number of physically distinct resources are likely due to different rankings of consumer fitness parameters in different patches.
This ranking must imply that each consumer species has some types of patches where it has the lowest R*. In general, neither of the mechanisms considered above leads to a distribution of consumers characterized by equal fitness in all patches. This outcome requires adaptive movement and is discussed in Section 10.5.10.4.2 Competition with mobile consumers and interference competition
All the models discussed above have assumed purely exploitative competition. If a consumer’s abundance has a direct effect on one or more of fitness parameters of other consumer individuals, this implies an ‘interference’ component to the competitive process. As noted in previous chapters, within a single patch interference can result in a priority effect, in which an initially more abundant consumer species is able to exclude an initially less abundant one. Alternative exclusion with a priority effect is the standard outcome within a single patch when there is only a single resource type, when attack rates (C) are negatively affected by consumer abundance, and when the magnitude of the effect on the attack rate parameter is greater for heterospecifics than it is for conspecifics. This leads to competition coefficients greater than unity. This may also occur with large interspecific effects on other fitness parameters.
The possibility of priority effects within at least some patches implies the potential for two or more system-wide attractors for metacommunities having alternative attractors in a single-patch setting. The simplest 2-competitor-2-patch system could have species 1 in both patches, species 2 in both patches, and two different configurations having a different species in each patch. The number of alternative outcomes increases with either more patches or more consumer species. Levin (1974) was apparently the first to note this fact. However, at least some of its implications seem to be largely ignored. The primary implication is that having higher levels of interspecific interference often makes coexistence more likely at the metacommunity scale, even though it makes coexistence within a patch less likely.
The presence of interspecific interference can be predicted on evolutionary grounds in some systems, and theywere observed in some of the earliest laboratory studies of competition between different species of the flour beetle genus, Tribolium (Park 1948). If the initial relative abundances of two such competitors differ in two patches, then it is likely that both will persist in the metacommunity, although each patch will consist almost entirely of one species.
The 2-species-2-patch system described in the previous paragraph has a different response of population sizes following a system-wide increase or decrease in one consumer’s mortality rate depending on the initial state of the system. It is simplest to describe these relationships using the Lotka-Volterra (LV) model for dynamics within a patch; in this model, alternative attractors within a patch can occur when the product of the two competition coefficients exceeds unity.
Consider a multi-patch system with two competitors and with alternative exclusion outcomes possible in some or all patches before the mortality perturbation. In the LV model this implies that the product of the two competition coefficients is > 1. It is necessary that each species of consumer be present somewhere in the system for changes in species composition to occur as one species experiences a mortality rate perturbation in all patches. Given that this condition is met, the equilibrium population size of the non-perturbed competitor will initially not be affected by increasing mortality of the focal species. However, when mortality on one consumer species produces a sufficiently low population size of that species in a given patch, invasion by the non-manipulated species will become possible, and its equilibrium will jump discontinuously to its carrying capacity within that patch. The focal species’ equilibrium in this patch will drop discontinuously to zero. These discontinuities are likely to occur at a different level of general mortality of the manipulated species in different patches because growth parameters of both species will usually differ across patches. The earliest extinctions occur in those patches where the initially present consumer had relatively low equilibrium abundance in the pre-perturbation system. There will then be a series of discontinuous jumps in system-wide equilibrium population size of the competitor as the manipulated species experiences greater mortality across all patches and is displaced successively in the patches where it has a lower ability to exclude its competitor. However, this relationship depends on the initial configuration of the system when the perturbation occurs; this configuration is determined by the initial abundances of consumers and resources in each patch. All these qualitative features are possible for consumer-resource models that also have alternative exclusion outcomes within patches.
10.5
More on the topic Random movement and coexistence:
- Random experiments
- What do coexistence and exclusion mean?
- Random variables
- The strength of competitive interactions affects coexistence
- Evolution and coexistence: current theory and the future
- Genetic drift results from random events
- Competitive Coexistence
- Mashkovskii and Tiagin: Random Selections or a Conscious Targeting of Scapegoats?
- We have seen in previous chapters that disturbance, stress, and predation can modify species interactions and allow for species coexistence.
- Early studies suggested that resource partitioning was the main mechanism of coexistence
- When ecologists perform experiments, they often take the three additional steps described in ECOLOGICAL TOOLKIT 1.1: they replicate each treatment, they assign treatments at random, and they analyze the results using statistical methods.
- Modern Finance, Efficient Market Hypothesis, Random Walk Hypothesis, and Empirical Evidence
- The Renewal Movement