The efficiency of energy transfer varies among consumers
Not all of the food energy consumed by a heterotroph gets incorporated into heterotroph biomass. Some is lost to respiration and excretion. We can use the concept of energy efficiency, defined as the output of energy per unit of energy input, to characterize the transfer of energy between trophic levels.
In studies of energy transfer in trophic systems, the concept of trophic efficiency is used, defined as the amount of energy at one trophic level divided by the amount of energy at the trophic level immediately below it. Trophic efficiency incorporates the proportion of available energy that is consumed (consumption efficiency), the proportion of ingested food that is assimilated by the consumer (assimilation efficiency), and the proportion of assimilated food that goes into producing new consumer biomass (production efficiency) (FIGURE 21.7).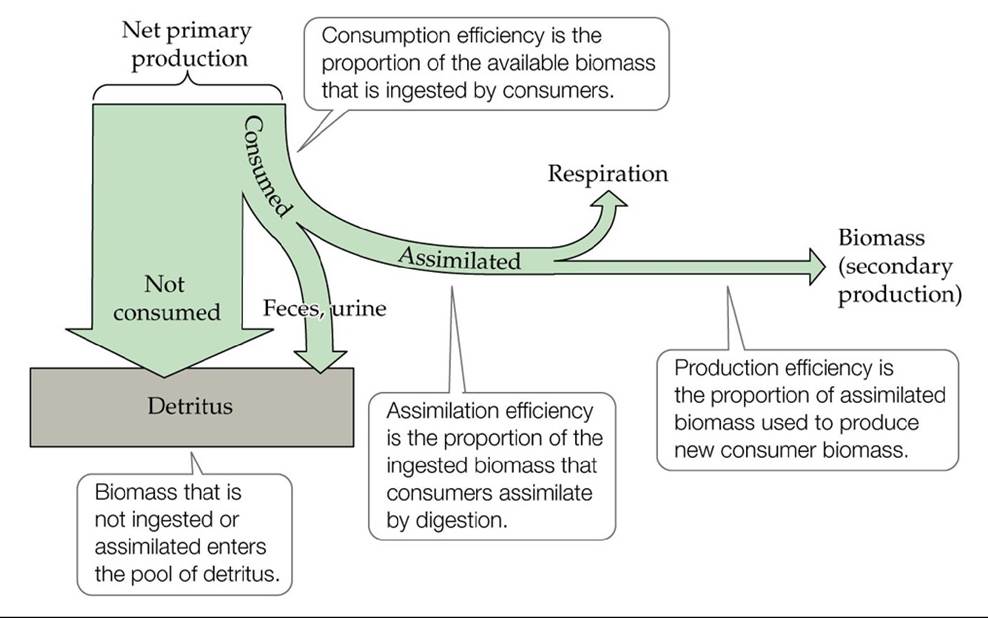
FIGURE 21.7 EnergyFlowandTrophicEfficiency Theproportionofenergytransferred between trophic levels depends on efficiencies of consumption, assimilation, and production.
How do the trends in consumption efficiency vary in Figure 21.6? What does this variation suggest about differences in consumption efficiency in aquatic versus terrestrial ecosystems?
(After F. S. Chapin et al. 2002. Principles of Terrestrial Ecosystem Ecology. Springer-Verlag: New York.) View larger image
As we saw above, not all of the biomass available at one trophic level is consumed by the next trophic level. The proportion of the available biomass that is ingested is the consumption efficiency. Consumption efficiency is typically higher in aquatic ecosystems than in terrestrial ecosystems (see Figure 21.6). Consumption efficiencies also tend to be higher for carnivores than for herbivores, although a systematic survey comparing the two groups has not been done.
Once biomass is ingested by the consumer, it must be assimilated by the
digestive system before the energy it contains can be used to produce new biomass. The proportion of the ingested food that is assimilated is the assimilation efficiency. Food that is ingested but not assimilated is lost to the environment as feces, entering the pool of detritus, or as urine. Assimilation efficiency is determined by the quality of the food (its chemical composition) and the physiology of the consumer.
The quality of the food available to herbivores and detritivores is generally lower than that of the food available to carnivores. Plants and detritus are composed of relatively complex carbon compounds, such as cellulose, lignins, and humic acids, that are not easily digested. In addition, plants and detritus have low concentrations of nutrients. Animal bodies, on the other hand, have carbon:nutrient ratios that are usually very similar to those of the animals consuming them and so are assimilated more readily. Assimilation efficiencies of herbivores and detritivores vary between 20% and 50%, while those of carnivores are about 80%.
How thoroughly food is digested is influenced by the consumer's thermal physiology and the complexity of the consumer's digestive system. Endotherms tend to digest food more completely than ectotherms, due to higher thermal stability and a tendency to have a more developed digestive system, and therefore have higher assimilation efficiencies. Additionally, some herbivores have mutualistic symbionts that help them digest cellulose. For example, as described in Concept 5.4, ruminants have a modified stomach chamber that contains bacteria and protists that increase the breakdown of cellulose-rich foods. This mutualistic symbiosis, coupled with a longer period of digestion, gives ruminants higher assimilation efficiencies than nonruminant herbivores.
Assimilated food can be used to produce new biomass in the form of consumer growth and production of new consumer individuals (reproduction).
However, a portion of the assimilated food must be used for respiration associated with maintenance of existing molecules and tissues as well as with construction of new biomass (see Concept 5.4). The proportion of the assimilated food that is used to produce new consumer biomass is production efficiency.Production efficiency is strongly related to the thermal physiology and size of the consumer. Endotherms allocate much of their assimilated food to metabolic production of heat and therefore have less energy left over to allocate to growth and reproduction than ectotherms do (TABLE 21.1). Thus, ectotherms have considerably higher production efficiencies than endotherms. Body size in endotherms is an important determinant of heat loss and thus of production efficiency. If body shape and insulation (fat, feathers, and fur) are held constant, then, as animal body size increases, the surface area-to-volume ratio decreases. Thus, a small endotherm, such as a shrew, will lose a greater proportion of its internally generated heat across its body surface than a large endotherm, such as a grizzly bear, and thus a small endotherm will tend to have a lower production efficiency than a large endotherm.
TABLE 21.1 ProductionEfficienciesofConsumers
| Consumer group | Production efficiency (%) |
| Endotherms | |
| Birds | 1.3 |
| Small mammals | 1.5 |
| Large mammals | 3.1 |
| Ectotherms | |
| Fishes and social insects | 9.8 |
| Nonsocial insects | |
| Herbivores | 38.8 |
| Detritivores | 47.0 |
| Carnivores | 55.6 |
| Non-insect invertebrates | |
| Herbivores | 20.8 |
| Detritivores | 36.2 |
| Carnivores | 27.6 |
Sources: F. S. Chapin et al. 2002. Principles OfTerrestrial Ecosystem Ecology. Springer-Verlag: New York; W. F. Humphreys. 1979. J Anim Ecol 48: 427-454.