NUCLEOTIDE REVERSE TRANSCRIPTASE INHIBITORS (NRTIs)
Each of the currently approved therapeutics in this class — abacavir, didanosine, emtricitabine, lamivudine, stavudine, zalcitabine, and zidovudine — exerts its activity after intracellular conversion to an active triphosphate form.
These active compounds reduce viral replication by competitively substituting for natural deoxynucleoside triphosphates, causing premature DNA-chain termination when incorporated. Human DNA polymerases generally remain unaffected by this class of compounds, although mitochondrial DNA polymerase-γ is an exception.4,5 Most long-term toxicity seen with this class of therapeutics is thought to result from mitochondrial DNA inhibition/depletion,6 possibly coupled with decreases in adenosine triphosphate (ATP) production or increases in reactive oxygen species generation.7 Supporting a connection between such toxicity and apoptosis, it has been shown that such disturbances are efficient to induce apoptosis in a variety of cells and cell lines upon exposure.Research into the potential cytotoxic properties of NRTIs began when (in an effort to explore the roots of muscular toxicity and mitochondrial myopathy seen in HIV patients) it was noted that histological evidence of mitochondrial damage correlated with long-term or heavy cumulative doses of zidovudine.8,9 Further studies by Jay et al. hinted at a dose-dependent phenomenon through the demonstration that low-dose-AZT-receiving patients lacked typical features of zidovudine myopathy.10 Depletion of muscle mitochondrial DNA was first noted in zidovudine (AZT) monotherapy patients,11 and this was coupled with results showing impaired cytochrome c oxidase activity, altered morphology,12 and altered oxidative phosphorylation enzymes in treated mitochondria.13 Shortly thereafter, it was demonstrated that these were not phenomena related simply to HIV infection, as such deficiencies were absent in HIV-positive patients not receiving AZT.14 Also, although this research did not directly demonstrate or assess drug-induced apoptosis, it provided a theoretical basis to investigate mitochondrial-dependent apoptosis modulation under these therapies.
Initial studies assessing apoptosis in short-term cultures of HIV-infected peripheral blood mononuclear cells (PBMCs) failed to suggest significant differences between AZT-treated and nontreated PBMCs,15 but confounders related to infection may have obscured examination of direct pharmacologic effects. AZT was later shown to induce biochemical and morphological changes characteristic of apoptosis in vitro using the mouse myeloma cell line Sp2∕O.16 Likewise, the treatment of noninfected, organotypic cocultures of spinal ganglia, spinal cord, and skeletal muscle from fetal rats with various concentrations of AZT also demonstrated duration-dependent effects on their morphology characteristic of apoptosis.17 Further studies by Viora et al. helped explain any previous conflict of research results by indicating that even though stimulated PBMCs seemed to be resistant to apoptosis in short-term culture, both zidovudine and zalcitabine (ddC) inhibited cell growth and cell cycling in CEM and phytohaemagglutinin (PHA)-stimulated PBMCs, and both compounds likewise induced apoptosis (in addition to growth and cycling inhibition) in CEM cell line cells.18 Of these two NTRIs examined, ddC seemed to be more efficient at reducing cell growth and inducing apoptosis, although a combination of the two seemed to display additive effects. This differential toxicity also matched the relative affinities of the two compounds for mitochondrial DNA polymerase-γ. Interestingly, limited research suggested that chronically HIV-infected T cell lines (MOLT-4/IIIB and CEM/ROD) are selectively more sensitive to apoptosis induced by stavudine than are their parental lines.19 Because this increased sensitivity to treatment is still presumed to be based on drug interference with DNA polymerase-γ, it hints at additional or enhanced vulnerabilities to mitochondrial damage potentially exploitable in infected cells.
What was more conclusively shown, in multiple large, prospective, open label, and randomized switch studies, is that stavudine therapy (and to a lesser extent, zidovudine therapy) is associated with greater patient risk for developing lipoatrophy.20,21 Lipoatrophy, which is a fat-wasting syndrome observed in some HIV patients, is recognized to occur most frequently in those patients taking certain NRTIs,22,23 although individuals receiving particular protease inhibitor therapies are also at risk for developing this syndrome.24,25 T.
Saint-Marc et al. have reported that the use of stavudine significantly correlates with fat wasting in both NRTI and protease inhibitor treatment groups, with odds ratios of 413 and 2.08, respectively, compared with zidovudine usage.26 Additionally, the severity of fat wasting seems to correlate well with the duration of NRTI therapy, which is further confirmed by direct observation of the correlation between mitochondrial DNA depletion and increased apoptotic cells in fat biopsies taken from treated patients.27Therefore, the majority of data available on the direct effects of NRTIs point to some limited ability of NRTIs to induce apoptosis in cells via their mitochondrial toxicity, at high concentrations or over long-term administration. This hypothesis can be examined by comparing the relative mitochondrial polymerase-γ inhibition versus apoptotic potential of currently available NRTIs. Ranking compounds for their ability to interfere with/inhibit DNA polymerase-γ was determined as (shown in decreasing order): zalcitabine, didanosine, stavudine, lamivudine, zidovudine, abacavir, and emtricitabine.28-32 What remains to be examined is a full account of the relative apoptotic potential across this class of compounds. To date, results have been suggestive that this correlation is tight.18,19,32 Additional evidence that mitochondrial toxicity may be the apoptotic mechanism functioning in NRTI therapy is suggested by noting common side effects of long-term therapy, which include peripheral neuropathy, myopathies, pancreatitis, and bone marrow suppression, that closely resemble certain heritable mitochondrial disease phenotypes.33,34 Although the exact mechanisms connecting NRTI-induced mitochondrial toxicity with resultant apoptosis remain unclear, several pathways are possible (see Figure 25.1) and suggested in the models below:
Mitochondrial DNA polymerase-γ interference: The interference of NRTIs with mitochondrial DNA (mtDNA) replication may result in increased mtDNA mutations, deletions, or simply competitive inhibition and continued replication prevention. All these could engender both enhanced vulnerability to and accumulation of additional mtDNA damage as well as directly impact the expression and function of enzymatic subunits encoded by the small mitochondrial genome.
Indirect impairment of mitochondrial enzymes through DNA damage (production, replacement problems) would lead to lowered ATP production35 and uncoupling of oxidative phosphorylation,36 finally triggering apoptosis.Electron transport chain or ATP production disruption: Alternately, NRTIs may directly impact the enzymes and processes of energy formation in mitochondria. This is suggested by one study that found that AZT can bind to and inhibit the enzyme adenylate kinase in vitro, resulting in a 30% decrease in ATP production at concentrations of 10 μM zidovudine.37 Although this concentration is unlikely to be physiological, other cell-culture and cell-free
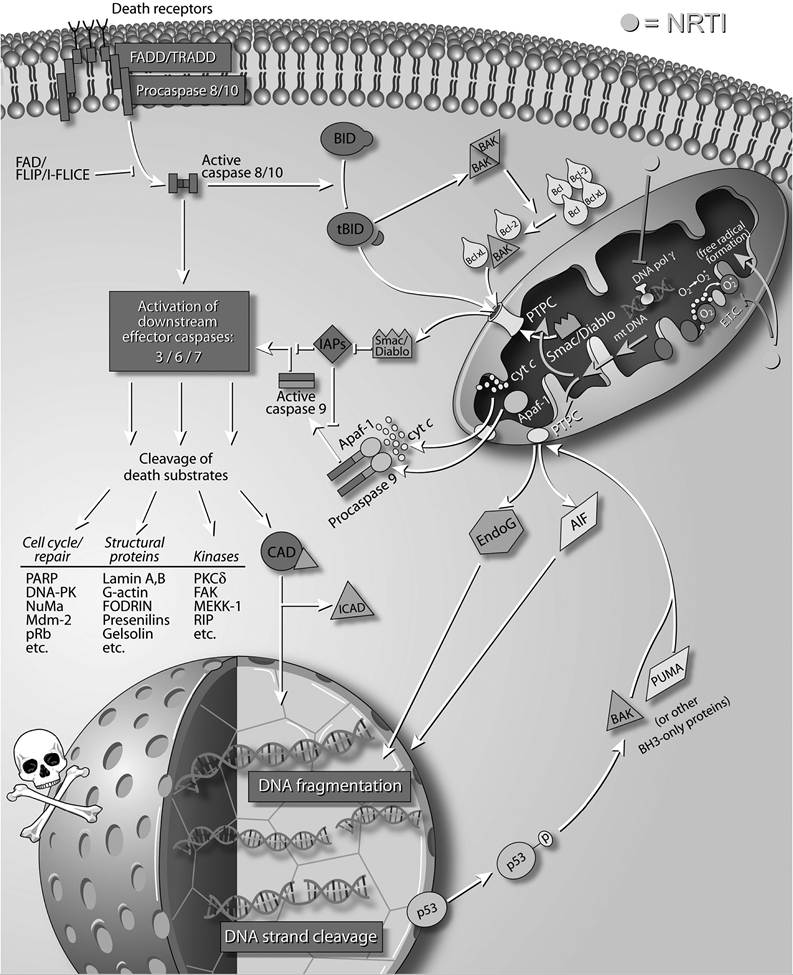
FIGURE 25.1 Apoptotic influences of Nucleoside Reverse Transcriptase Inhibitors (NRTIs). As described in the text, NRTIs likely impact cellular apoptosis through a number of potential mitochondrial interactions. Interference with mitochondrial DNA polymerase gamma, disruption of the electron transport chain (and ATP production), and enhanced reactive radical formation by this class of HIV therapeutics are all proposed to feed forward toward mitochondrial deregulation, which results in the release of pro-apoptotic factors.
studies have shown that AZT also inhibits the adenosine diphosphate (ADP)Zadenosine triphosphate (ATP) translocator38 and demonstrates a dose-dependent inhibition of both cytochrome c reductase and NADH-Iinked respiration.39
Reactive radical formation: Finally, NRTIs may exert their toxic effects through the generation of oxygen-free radicals that attack mtDNA and induce mutation.40 This hypothesis was supported by two studies in which zidovudine-treated mice were killed and their liver levels of oxidized guanosine were determined as a measure of oxidant damage. In both studies, greater levels of oxidized guanosine were found in the AZT-treated mice than in controls, and the administration of antioxidants inhibited this effect.40,41 Although the pathway of NRTI-promoted radical formation is lacking, in this scenario damage to the mtDNA results from oxidation-induced mutation rather than from substitution-induced termination or misincorporation.
In the interest of preserving perspective, it deserves mention that although there are studies indicating the proapoptotic effects of NRTIs, this is due in part to the success of NRTIs as therapeutics, and the careful clinical examinations of NRTI toxicity limits. When used in practice for the management of HIV infection, multiple drugs are often prescribed together, or therapies are alternated, in a manner that intentionally reduces the incidence or severities of drug toxicities. The result of administration within the therapeutic index for NRTIs has been resoundingly beneficial for patients, and it is certain that in the context of HIV infection, wherein large numbers of uninfected “bystander” T cells are likely to be undergoing apoptosis, administration of NRTIs to these patients reduces their spontaneous lymphocytic apoptosis levels.42 This reduction in apoptosis generally mirrors a concurrent suppression of viral load and is, therefore, believed to be an indirect effect mediated through a reduction in the quantity of proapoptotic viral proteins and virions. However, there was a recent report suggesting a potential antiapoptotic effect attributable to a model system for AZT, observed in treatment of maedi-visna-virus-infected sheep choroid plexus cells.43 Researchers observed the inhibition of morphological characteristics, as well as inhibition of caspase-3, -8, and -9 cascades, DNA laddering, and mitochondrial apoptosis-inducing factor (AIF) leakage, although whether this represents a direct protective effect of AZT or, more likely, an indirect result of the reduction in viral products and their downstream effects remains to be seen. Further research is needed, but the blockade of both caspase-dependent and caspase-independent pathways suggested in this report is intriguing.
There is another structurally distinct class of NRTIs that differs from nucleoside inhibitors in that they possess a phosphonyl moiety that makes these compounds resistant to enzymatic hydrolysis.
Prototypes for this class of drug include tenofovir disoproxil fumarate and adefovir dipivoxil, and these compounds are able to circumvent the obligatory intracellular phosphorylation step of nucleoside inhibitors before being pharmacologically active. Unfortunately, they also demonstrate some inhibitory activity for DNA polymerase-α, -δ, and -ε, which makes them less specific for viral inhibition and reciprocally more toxic.44 Current research on this class of compounds demonstrates that they can be very efficient at inducing differentiation of in vitro tumor-cell models, possess antitumor activity in vivo, and cause apoptosis in a wide variety of cell lines.44-46Although clear experiments are lacking, it is likely that such compounds induce apoptosis through the same mechanism(s) as NRTIs. In both a nasopharyngeal carcinoma tumor xenograft mouse model (in which EBV is present) and in MTT assays of the EBV-positive epithelial cell line NPC-KT, to apoptosis as indicated by DNA fragmentation assays and evidence of poly-ADP ribose polymerase (PARP) cleavage, was the result of treatment with micromolar amounts of several phosphonated nucleotide analogues.47 The observed apoptotic effects in these experiments are believed to be independent of EBV-polymerase inhibition, as that protein is not expressed in latently infected NPCs.48 The basis for apoptotic activity in this class of compounds is their broader effects on various cellular DNA polymerases. Impacting and decreasing nuclear DNA as well as mitochondrial DNA reasonably exacerbate the toxicities described above for mitochondrial DNA polymerase interference. Although only two nucleotide compounds are currently approved for HIV therapy, there are likely to be further refinements and advances in these phosphonated compounds, and with them will come further elucidation of their abilities to induce apoptosis.
More on the topic NUCLEOTIDE REVERSE TRANSCRIPTASE INHIBITORS (NRTIs):
- NUCLEOTIDE REVERSE TRANSCRIPTASE INHIBITORS (NRTIs)
- NONNUCLEOSIDE REVERSE TRANSCRIPTASE INHIBITORS (NNRTIs)
- COMPLICATIONS OF THERAPY WITH NUCLEOSIDE ANALOGUE REVERSE TRANSCRIPTASE INHIBITORS
- Appendix 2 Interactions Between Antiretrovirals and Drugs Commonly Used to Treat Cardiovascular Disease
- Chloroform Toxicity
- FUSION INHIBITORS
- HIV Type 1
- The Legacy of the Aryan Pastoralists
- DETECTION OF DNA FRAGMENTATION
- CLASS I IFNs