Central nervous system cytology
Cytology of CNS parenchyma is becoming increasingly utilized in veterinary medicine, as it can be used to good effect in conjunction with imaging to diagnose and plan treatment in patients with CNS lesions (Powell, 2005).
Fine needle aspiration (FNA) is an excellent modality for deep-seated tumors where direct visual access would require disruption of a large amount of normal nervous tissue. Although the acquisition of samples and the practice of CNS cytology are highly specialized, a basic overview is included to help guide diagnostic procedures and patient care.Sampling
CNS biopsy can be obtained through less invasive ultrasound or CT-guided (either free-hand or stereotactic) FNAs or by more invasive open craniotomy accessed FNA or incisional biopsy techniques (Platt et al., 2002). The exact procedure for acquiring these samples is complex, necessitates specialized imaging and equipment, and is outside the scope of this text. Consultation with a neurologist or surgeon with experience in CNS cytologic collection is recommended.
The basic principles that need to be observed include preparation of a fresh sample that is thin enough for cytologic evaluation.
CNS cells deteriorate quite rapidly, causing a loss of cellular structure and differential staining. Sample processing is often expedited for intraoperative samples by including the cytologist in scheduling the surgery and staining samples quickly. Samples collected post-mortem need special care. Although a delay of hours is acceptable for histologic evaluation, it will likely significantly affect cytologic evaluation and should be avoided.
Sample thickness is another concern. Much of the CNS lacks significant fibrous connective tissue in health. This makes it very soft and makes a squash type preparation of material collected by FNA, at surgery, or at gross necropsy very easy.
One exception to this rule is a sample with large numbers of glial cells; these can be more challenging to spread and produce an even distribution of cells (Lee & Tihan, 2015). Less than a cubic millimeter of tissue is needed for a high-quality squash preparation. Multiple thin smears are preferred to a single dense sample. Touch imprints of samples collected for histopathology can be used to produce an interpretation (Powell, 2005).Several staining techniques can be used for CNS cytology. Alcohol fixed wet slides can be stained with hematoxylin and eosin (Iqbal et al., 2006). Although this is not a commonly used cytologic stain, it has the main advantage of being quick. The sample is alcohol fixed when still wet and immediately stained; this allows a diagnosis within 10 minutes of sample collection in some cases. Romanowsky-type stains can be used on dried slides and are more commonly utilized.
Indications and contraindications
The same basic principles outlined for CSF collection hold true for indications and contraindications of CNS cytology. One obvious caveat is that the lesions must be physically within the reach of the cytology needle or the surgeon’s scalpel. Additionally, important, uninvolved structures cannot lie along the path of needle insertion or significant complications can arise. Tumor seeding along the sampling tract has been reported in human medicine; the occurrence of iatrogenic tumor spread in veterinary medicine is unknown (Patrick et al., 2006).
One common use for intraoperative cytologic evaluation of CNS samples is to determine the diagnostic adequacy of a sample collected for histologic evaluation. In human medicine it has been noted that only one-third of the first samples collected by stereotactic techniques are diagnostic. A quick review of a smear can indicate that more samples are needed to reach a diagnosis before the patient leaves the operating room (Lee & Tihan, 2015).
Cytologic evaluation of a series of ten canine brain tumors with concurrent histologic evaluation found that the two modalities shared only 50% agreement for a specific diagnosis, although all FNA samples were confirmatory for neoplasia (Platt et al., 2002).
In human medicine, where the technique is used much more frequently, a lack of agreement is only found in 3% of cases (Lee & Tihan, 2015). Given the diagnostic benefits, the economic cost, and the minimally invasive nature of CNS FNA cytology, even the authors that found a poor agreement between cytology and histopathology still advocate FNA of intracranial lesions. Additionally, cytologic evaluation of samples obtained at necropsy can be a quick and inexpensive way to identify underlying pathology. A better description of the pathologic process that led to the demise of a beloved pet can sometimes help in the acceptance of pet loss.Normal cytologic findings
The CNS is composed of a multitude of cells, each with their own specialized function and form. Within the category of neuroepithelial cells are neurons and glial cells. The glia can be further stratified into astrocytes, oligodendrocytes, Schwann cells, ependymal cells, and choroid plexus cells. Neuroepithelial cells derive from the neuroectoderm. CNS cells derived from the mesenchyme include meningeal cells and microglia.
Neuropil
Neuropil is a meshwork of axons, dendrites, synaptic junctions, and glial fibrillary processes. As such, it is made of parts of neurons, astrocytes, oligodendrocytes, microglial cell membranes, and extracellular matrix (Miller & Zachary, 2017). Cytologically, neuropil appears as a fine eosinophilic felt-like meshwork interwoven with nuclei from neurons and glial cells (Figures 14.26a–d).
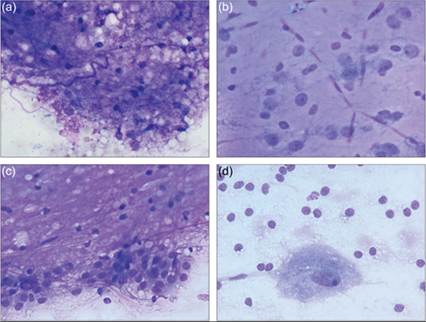
Figures 14.26a–d Squash preparations of normal CNS tissue in dogs. (a) A frothy pink to purple extracellular material with interspersed small, condensed nuclei on CSF analysis is consistent with neuropil. Note how both diffuse material and long ribbon-like structures are present (Wright–Giemsa, 500? magnification). (b) A preparation of gray matter from the cerebral cortex shows many fine capillaries filled with red blood cells.
The smallest, condensed nuclei are most consistent with oligodendrocytes. These cells have a large amount of cytoplasm; however, it is found distant to the nuclei, wrapped around neuronal processes. The nuclei located next to the vasculature, with open chromatin and moderate poorly discernible basophilic cytoplasm, are most consistent with astrocytes. The larger nuclei with a distinct nucleolus and more abundant cytoplasm belong to neurons (Wright–Giemsa, 1,000? magnification). (c) A preparation of white matter from the cerebral cortex has more prominent felt-like neuropil. The more condensed round nuclei causing nonstaining defects in the neuropil are most consistent with oligodendrocytes. In this location astrocytes are classified as fibrous astrocytes; they have smaller nuclei and sparsely branched processes. Significant numbers of neuron cell bodies are not expected in the white matter; however, neuronal dendrites and axons contribute to the felt-like network in the neuropil. The cuboidal to columnar cells along the bottom of the image are cytologically consistent with ependymal or choroid plexus cells; in this location they are expected to be ependymal in origin (Wright–Giemsa, 1,000? magnification). (d) An imprint from canine cerebellum. The single large neuron in the lower half of the image has abundant amounts of granular basophilic cytoplasm, few clear cytoplasmic vacuoles, and a large ovoid nucleus with stippled chromatin and single large nucleolus. Given the location, this large neuron is a Purkinje cell and the smaller round cells in the image are likely a combination of glial cells and neurons from the inner granular layer. The background consists of wispy fibrillar cell processes from neurons and glial cells (Wright–Giemsa, 1,000? magnification).
Neurons
Larger neurons will often have a triangular shape, ample amounts of basophilic, somewhat granular cytoplasm, a large nucleus, and a large prominent nucleolus (Figures 14.26a–d, 14.27; Lee & Tihan, 2015).
The long cytoplasmic processes of the neuron will often produce a pink, amorphous to faintly fibrillar appearance surrounding the neuron. This fibrillary material can also be seen arrayed around glial cells as part of their glial fibrillary processes. The inner granular layer of the cerebellum is composed of much smaller neurons, which produce a population of small round basophilic cells. These cells can be mistaken for glial cells if the location of the sample site is not known or if the significance of interspersed large neurons, such as Purkinje cells, is overlooked (Lee & Tihan, 2015). 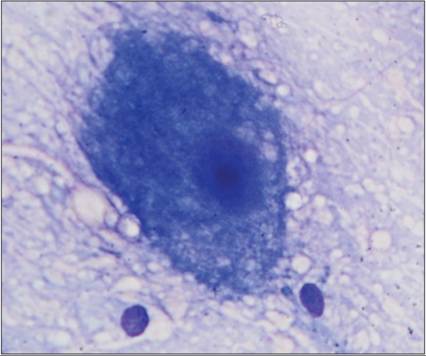
Figure 14.27 A large neuron with abundant granular basophilic cytoplasm, large nucleus, and prominent nucleolus is present in an imprint of gray matter from the cerebral cortex (Wright–Giemsa, 1,000? magnification).
Astrocytes
Astrocytes are glial cells that form the blood–brain barrier and participate in repair and support of the CNS (Figures 14.26a–d). To do this they maintain foot processes along the blood vessel and surrounding axon. Astrocytes make up the predominant cell type in the CNS (Miller & Zachary, 2017).
They are basophilic cells smaller than large neurons and have open chromatin on cytology; their finer processes are not commonly appreciated. Histologically, they are divided into protoplasmic astrocytes which have short cellular processes and fibrous astrocytes which have long branching processes. The protoplasmic astrocytes are found within the gray matter while fibrous astrocytes are found within the white matter.
Oligodendrocytes
Oligodendrocytes play a role in myelination of axons (Figures 14.26a–d). To do this they have a large amount of cytoplasm, which extends out in thin processes and wraps around nearby neurons. A single oligodendrocyte can interact with up to 50 neurons. However, most of this cytoplasm is not directly around the nucleus, leading to frequent confusion of oligodendrocyte nuclei and lymphocytes.
Microglia
Microglia are thought to be the CNS resident component of the monocyte–macrophage system. Their primary role is the removal of cellular debris and fluid via phagocytosis and pinocytosis. Interestingly, although microglia are the resident monocyte–macrophage population, up to 70% of the macrophages in an inflammatory or degenerative response within the CNS are derived from blood monocytes. Although several histologic forms occur, including amoeboid and ramified microglia, on cytologic evaluation they are typically smaller cells with round to rod-shaped nuclei and scant cytoplasm. Microglial cells appear similar to macrophages located elsewhere, with foamy, lipid filled cytoplasm (Figure 14.28).
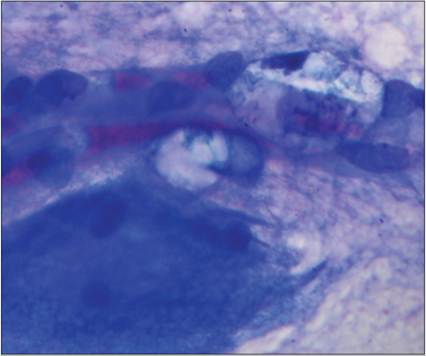
Figure 14.28 A red blood cell filled vessel runs horizontally through the image. Two microglial cells are associated with the vessel. They have foamy clear cytoplasm and rod-like nuclei. A neuron cell body is present in the lower left of the image (Wright–Giemsa, 1,000? magnification).
Meningeal cells
There are three layers in the meninges: the dura mater, arachnoid membrane, and pia mater (Miller & Zachary, 2017). The pia mater is closely adhered to the brain and spinal cord. It is a layer of fibroblasts that can be as thin as a single cell layer. The pia mater forms fibrous connections with the arachnoid membrane. The arachnoid membrane is a multilayered mesenchymal tissue. Nerve bundles and blood vessels are found in the space between the arachnoid membrane and the pia mater. This subarachnoid space is also the location most commonly accessed for collection of CSF. Together the arachnoid and pia mater are referred to as the leptomeninges. Lining the outer surface of the arachnoid membrane and inner surface of the dura mater is a single layer of mesothelial-like cells whose function is similar to mesothelium in the abdomen and thorax; it allows frictionless movement of the brain and spinal cord within the bony vault of the calvarium and spinal canal. The dura mater is a dense collagenous membrane. It is bilayered and acts as the inner periosteal layer of the calvarium and spinal canal. The bilayer splits in the spinal canal and creates the extradural space.
Cytologically, meningeal cells can be found in sheets or individually; they have moderate amounts of faintly eosinophilic cytoplasm and nuclei that are variably round to ovoid to elongate (Figures 14.29a, b). The cytoplasmic borders are often ill defined; however, storiform or pseudoacinar arrangements can be seen.
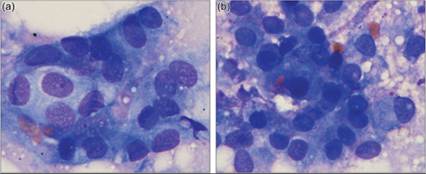
Figures 14.29a,b Imprint of the dura. (a) Meningeal cells are present in a sheet of polygonal to spindloid cells with poorly distinct cell borders and round to oval nuclei. (b) A pseudoacinar arrangement is present in the center bottom of the field (Wright–Giemsa, 1,000? magnification).
Ependymal cells
Ependymal cells are cuboidal to columnar epithelial cells that have basally located nuclei and apical cilia (Miller & Zachary, 2017). Their primary function is the movement of CSF through the ventricular system. Ependymal cells are joined by gap junctions. This allows transfer of proteins and fluid from the CSF, around the ependymal cells and into the extracellular spaces of the brain. Ependymal cells are found in the ventricles, the mesencephalic aqueduct, and the central canal of the spinal cord. Cytologically, it is challenging to distinguish ependymal cells from choroid plexus cells (Figure 14.26c).
Choroid plexus cells
In situ, choroid plexus cells are specialized ependymal cells arranged in fronds. They produce approximately 90% of the CSF volume. Choroid plexus cells are cuboidal epithelial cells that are distinct from other ependymal cells. Among other features, choroid plexus cells have apical microvilli while ependymal cells are ciliated.
An excellent review of choroid plexus cells in multiple species has been published (Garma-Aviña, 2004). This study found four subtypes of choroid plexus cells. In the dog, most (~75%) choroid plexus cells are large polygonal cells measuring 15–20 μm. These cells are termed alpha cells and are characterized by countless basophilic granules and small, round highly regular nuclei lacking obvious nucleoli (Figure 14.30a). Approximately 25% of the cells are similarly sized to alpha cells but have gray to faintly acidophilic granules and are called beta cells (Figures 14.30b, c). Sometimes, a few tiny round clear vacuoles can be noted in beta cells. Their nuclei are slightly larger and paler with finer chromatin than alpha cells. Both alpha and beta cells are positive for pancytokeratin. A third type of cell, the gamma cell, is characterized by the presence of large membrane bound cytoplasmic vesicles that measure between 3 and 30 μm (Figure 14.30d). Pancytokeratin can stain fine linear structures within these vesicles; however, the majority of the cell is pancytokeratin negative. This cell type makes up about 1–3% of cells in the dog. Less than 1% of cells in normal canine choroid plexus are epiplexus macrophages, also called Kolmer cells. These large cells are interposed between the other cell types and have copious pale cytoplasm. Nuclei can be round, oval, or bi-lobed and the cytoplasm often contains fine clear vacuoles.
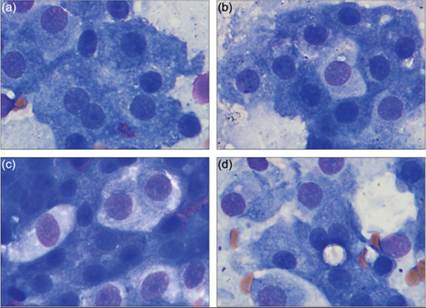
Figures 14.30a–d Cytologic imprints of the choroid plexus. (a) Seventy-five percent of the cells are polygonal cells with basophilic granules and small round nuclei. These have been called alpha cells. (b) A beta cell is seen in the center of the image; it is roughly the same size as the surrounding alpha cells and has a slightly larger, more open nucleus and eosinophilic to gray cytoplasmic granules. (c) Four beta cells are present in a dense cluster of alpha cells. (d) Very rarely, cells with a large membrane bound vesicle can be found in the choroid plexus. These are called gamma cells. Gamma cell vesicles were found extracellularly in the preparation (not shown) (Wright–Giemsa, 1,000? magnification).
More on the topic Central nervous system cytology:
- Barger A.M., MacNeill A.L. (Eds.). Small Animal Cytologic Diagnosis: Canine and Feline Disease. CRC Press,2024. — 536 p., 2024
- 44 Diseases of the Vulva and Vagina
- 30 Chronic Pelvic Pain
- Index
- Index