CHAPTER 41 HARMFUL ALGAL BLOOMS INCLUDING Cyanobacterial toxicosis
KJELL HANDELAND1 AND DOLORES GAVIER-WIDEN2
1Department of Animal Health, Section of Wildlife Diseases, National Veterinary Institute, Oslo, Norway 2National Veterinary Institute (SVA), and Swedish University of Agricultural Sciences, Uppsala, Sweden
INTRODUCTION
A number of aquatic microalgal species are known to cause harmful toxic blooms and animal intoxications (phycotoxicosis).
These include a variety of cyanobacteria (bluegreen algae) as well as dinoflagellate and diatom species. Although dinoflagellates have been associated with seabird mortalities, e.g. along the English coast(1), cyanobacteria are by far the most common group associated with animal toxicosis. Despite their aquatic origin, cyanotoxins appear to be more hazardous to terrestrial animals than to aquatic biota(2).Toxic cyanobacterial blooms have long been associated with intoxication events in domestic animals all over the world, and they are also posing a human health hazard (e.g.(3)). Some reports mention that wild mammals and birds were also affected. Specific reports of suspected cyanotoxic die-offs in wildlife are, however, mostly restricted to mass mortality events in birds(4’5). Owing to the accidental nature of exposure and limitations of diagnostic methodology, the general frequency of cyanotoxicosis in wildlife is likely to have been greatly underestimated in the past.
AETIOLOGY
Cyanobacteria (Class Cyanophyceae) are prokaryotic (no nucleus) photosynthetic microorganisms containing chlorophyll-a(6). They also contain accessory photosynthetic pigments such as phycocyanin, allophycocyanin and phycoerythrin. Cyanobacteria are important primary producers that may flourish in fresh, brackish and marine water ecosystems. Growth is dependent on carbon dioxide, inorganic compounds and light, and may take place both in the pelagic (water column) and benthic (sediment) water domains.
Their basic morphology is very diverse, including unicellular, colonial and multicellular filamentous forms. Cell colour varies from blue-green to violet- red, depending on the composition of photosynthetic pigments. Around 2,000 species of cyanobacteria, belonging to 150 genera, have been described, but only 40 species are associated with animal toxicoses.Cyanobacterial toxins constitute a diverse group of compounds from both a chemical and toxicological point of view(2). They can be classified according to their mode of toxicity into three different groups:
1. hepatotoxins, including microcystins and nodularins
2. neurotoxins, including anatoxins and saxitoxins
3. dermatotoxins.
Microcystins include 80 structural variants of cyclic peptides, among which microcystin- RR and -YR are recognized as the most toxic. Microcystins are produced by a relatively wide range of fresh and brackish water cyanobacteria, the most important belonging to the genera M icro- cystis, Anabaena, Planktothrix and Oscillatoria. Production of nodularins is restricted to the species Nodularia spumi- gena, which occurs in brackish waters and is an important species in the Baltic Sea.
Infectious Diseases of Wild Mammals and Birds in Europe, First Edition. Edited by Dolores Gavier-Widen, J. Paul Duff, and Anna Meredith. © 2012 Blackwell Publishing Ltd. Published 2012 by Blackwell Publishing Ltd.
The anatoxins involved in animal cyanotoxicoses are anatoxin-a, homoanatoxin-a and anatoxin-a(s). The anatoxins are mainly produced by cyanobacteria within the genus Anabaena, but also by other genera, including Microcystis, Oscillatoria and Planktothrix.
The dinoflagellate Karenia brevis produces brevitoxin, a potent marine neurotoxin.
EPIDEMIOLOGY
GEOGRAPHICAL DISTRIBUTION AND HOSTS
Most cases of cyano-intoxication associated with cyanobac- terial blooms have been reported from livestock and dogs. In Europe, domestic animal intoxication has been reported in Germany, Denmark, Sweden, Norway, England, Scotland, Ireland and Switzerland (e.g.(3)).
Although associated with deaths in various wild mammal species on other con- tinents(7), cyano-intoxication in wild European mammals seems to be restricted to a case report in roe deer ( Capreolus capreolus} in Norway(8). This diagnosis was based on demonstration of typical liver lesions and the presence of high concentrations of microcystin-YR in the liver tissue. The suspected source of intoxication was algal growth in a drainage ditch.Mortalities in wild waterfowl associated with cyanobac- terial bloom have been registered in several European countries including Denmark(9,10); Belgium(11), Spain(12’13), Sweden and Finland1-5). The Danish outbreaks were associated with anatoxin- a(s) producing A nabaena blooms in freshwater lakes. The Spanish outbreaks occurred in the Donana National Park, which is a major waterfowl refuge in Southern Europe. Here, many mass mortality events have occurred since 1973, and at least two were associated with heavy cyanobacterial blooms. In the first recorded outbreak, a large number of waterfowl, especially greater flamingos (Phoenicopterus ruber), died(5). This outbreak was associated with a mixed bloom, consisting of Microcystis aeruginosa and Anabaena flos-aquae. Laboratory examination revealed hepatotoxicosis and high levels of microcystins in the liver of dead birds. The second outbreak was associated with a bloom of Microcystis aeruginosa, and at least 6,000 waterfowl belonging to 47 species were killed(13). This outbreak initially affected herbivorous waterfowl and fish, followed by a die- off in piscivorous birds. Laboratory findings included typical liver lesions in dead birds, and the presence of high concentrations of microcystins in the livers of both dead birds and fish. The authors suggested this cascade of waterfowl die-offs could be explained by accumulation of cyanotoxins in the food web: firstly affecting herbivorous avian species consuming cyanobacterial bloom water (scum), and subsequent poisoning of piscivorous birds feeding on fish that had bioaccumulated the toxins through predation on contaminated zooplankton and aquatic invertebrates.
Bioaccumulation of cyanotoxins in the food web (mussels, fish, waterfowl) has also been demonstrated for nodularins produced by Nodularia spumigena in the Baltic Sea(14).Episodes of mortality associated with brevetoxicosis affected endangered Florida manatees ( Trichechus manatus latirostris) and bottlenose dolphins ( Tursiops truncatus) that had consumed toxic seagrass and fish, respectively1-15). Southern sea otters (Enhydra lutris nereis), a threatened species, were reported to have died of cyanotoxicosis in the Pacific coast of California1-16). Mortality of marine mammals caused by phycotoxicosis has not been reported in Europe.
ENVIRONMENTAL FACTORS
Planktonic (pelagic) cyanobacterial blooms are characterized by water discoloration and formation of a blue or green surface scum. Blooms occur in eutrophic waters rich in mineral nutrients (phosphorous, nitrate) during summer or autumn, when water temperature is high and light conditions favourable for photosynthesis. Cyanobacteria are slow-growing compared with other competing planktonic microalgae, and require stagnant water to proliferate in large numbers. They have, however, the advantage of needing less light (accessory photosynthetic pigments) than other microalgae, allowing them to outcompete other species under turbid water conditions. Under stagnant and turbid water conditions one or a few species of cyanobacteria may completely dominate, and if the dominating species include one or several toxin-producing cyanobacteria, intoxication events may occur (Figure 41.1). Toxic cyanobacterial blooms occur mostly in freshwater ecosystems (lakes, ponds, rivers) and have been reported from a large number of European countries(17). In Europe, some 53% of fresh water lakes are considered eutrophic and thus constitute potential sites of toxic blooms(18). Brackish (marine) ecosystems may also be eutrophic and develop
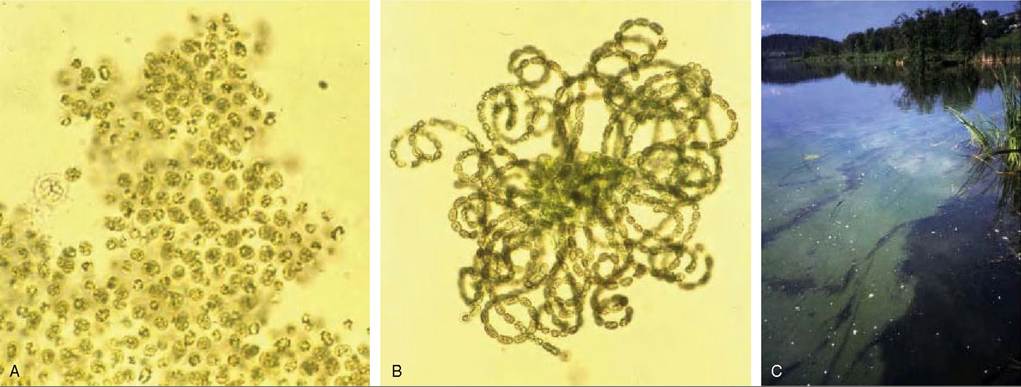
FIGURES 41.1 Mixed cyanobacterial bloom of Microcystis aeruginosa (A) and Anabaena flos-aquae (B) occurring in lake Ostensjovannet (C), Oslo, Norway in July 1989.
The bloom was associated with mass mortalities in ducks, geese and gulls. Some birds showed microscopic liver lesions consistent with hepatotoxicosis, whereas others showed less distinct liver lesions. Thus, the bird die-off may have been associated with both hepa- totoxic and neurotoxic cyanotoxins. Photos: Olav M. Skulberg, Norwegian Institute of Water Research.algal blooms, e.g. the Baltic Sea(6). Although some water sources are naturally eutrophic, in many others the excess nutrient levels are caused by human activity, resulting from wastewater discharges or run-off from agricultural areas.
Animals are most often exposed to toxic amounts of algae as a result of drinking water or scum from a lake or pond in which prevailing winds have concentrated the bloom. Cyano-intoxication may also occur following water consumption from smaller water sources such as waterholes, small dams and dugouts containing stagnant water, in which thick coats of algae may form(19). There are indications that animals do not avoid cyanobacterium- contaminated drinking water(20).
Intoxication may also be associated with benthic aggregations (coherent mats) of cyanobacteria. During periods of high photosynthesis, production of oxygen bubbles may increase the buoyancy of the mats, causing them to tear loose and rise to the surface, where if ingested, they may cause animal intoxication. This type of cyanotoxicosis has been reported in domestic animals in Scotland(21) and Switzerland(22). Another route of intoxication is through consumption of organisms (fish, molluscs, zooplankton) that have themselves fed on cyanobacteria or otherwise bio-accumulated cyanotoxins(2). In this way, mortality can occur weeks after the disappearance of the bloom.
PATHOGENESIS AND PATHOLOGY
Cyanobacterial toxins are released following algal cell destruction in the animal’s stomach, but may also exist free in the water body as a result of cyanobacterial cell autolysis.
Microcystins (and nodularins) are rapidly absorbed in the intestinal tract and transported to the liver, which constitutes the target organ(23). It has also been demonstrated that microcystins may be efficiently absorbed via the respiratory route. In the liver, microcystins and nodu- larins function as potent inhibitors of protein phosphatases 1 and 2A, leading to actin filament alterations and disruption of the hepatocyte cytoskeleton, as well as damage to the sinusoidal endothelium(24).The toxicokinetics of the neurotoxic anatotoxins have not been described. However, based on the peracute clinical course of this intoxication, intestinal absorption must be rapid. Anatoxin- a and homoanatoxin- a act as potent cholinergic agonists at nicotinic acetylcholine receptors in neurons and neuromuscular junctions, leading to respiratory paralysis. Anatoxin- a(s) is an irreversible acetylcholinesterase inhibitor, which leads to increased concentrations of acetylcholine in the synapse, resulting in neuromuscular blockage and respiratory arrest. The anatoxins do not cause cell-morphological changes.
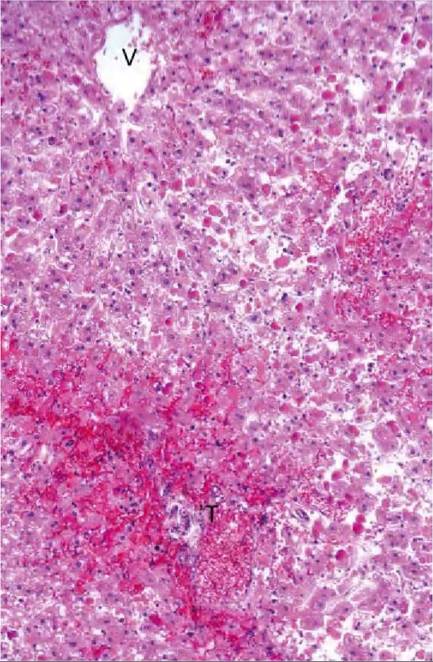
FIGURE 41.2 Liver from a microcystin- intoxicated roe deer, showing diffuse hepatocyte dissociation, degeneration and necrosis and perisi- nusoidalhaemorrhage.T = portal tract; V = central vein. Haematoxylin- eosin stain. From Handeland & 0stensvik, 2010(8).
Animals found dead after drinking toxic bloom may show evidence of green- or blue-coloured cyanobacteria on the hair or feathers. Gross pathological findings reported following acute microcystin and nodularin intoxication include a pale, swollen and sometimes haemorrhagic liver, as well as ascites and widespread small haemorrhages. Microscopic liver lesions include hepatocellular dissociation, degeneration and necrosis, as well as areas of perisi- nusoidal haemorrhage or massive bleedings (Figure 41.2). No specific gross or microscopic lesions are found in animals dying from anatoxin intoxications.
CLINICAL SIGNS
Clinical signs of cyanotoxicosis may be linked to either hepatotoxic or neurotoxic effects. However, it is important to keep in mind that mixed cyanobacterial blooms, with multiple associated toxins, are common, and may complicate the clinical picture. Moreover, the effect of interactions between the different toxins involved is not clearly understood. Thus, the clinical signs resulting from naturally occurring intoxication events may be complex.
The clinical course of fatal microcystin and nodularin hepatotoxicoses is acute, with death normally occurring within a few hours post- exposure. Clinical signs include general weakness, vomiting, diarrhoea, pallor and shock. Animals living beyond a few hours post-exposure may develop hyperkalaemia, hypoglycaemia, nervousness, recumbancy and convulsions.
The course of fatal neurotoxicoses caused by anatoxin-a, homoanatoxin-a and anatoxin-a(s) is peracute, with death due to respiratory paralysis. Death normally occurs within a few minutes to a few hours post-exposure. Clinical signs following anatoxin-a and homoanatoxin-a ingestion include rigidity, muscle tremor, cyanosis and paralysis and, in birds, opisthotonos. Clinical signs following anatoxin- a(s) intoxication include tremor, inco-ordination, and convulsion, as well as excess salivation (‘s’ stands for salivation), lacrimation and urinary incontinence.
DIAGNOSIS
Specific diagnosis of cyanobacterial toxicosis has been difficult, as a result of the lack of appropriate diagnostic procedures, and remains challenging. In the past, diagnosis was based upon algal bloom identification and disease history, possibly in combination with toxin analyses of algae and bioassays in experimental animals. Toxin production is, however, not constant during a bloom, and many algal blooms, even when dominated by species known to be capable of producing toxicosis, may not be hazardous.
A definitive diagnosis of cyanotoxicosis should be based upon a thorough post mortem examination, followed by demonstration of high tissue cyanotoxin levels. The post mortem examination should aim to demonstrate the presence of typical liver lesions by cyanobacterial hepato- toxicosis or absence of morphological lesions by cyanobacterial neurotoxicosis, as well as consideration of other possible aetiologies. One important differential diagnosis in avian events is botulism, a common cause of waterfowl death in eutrophic waters. Methodologies for the determination of cyanotoxins in post mortem tissues include liquid chromatography and mass Spectrometric systems, as well as enzymatic assays and immunoassays1-3). Liquid chromatography and mass spectrometric methods have been employed on tissue samples for quantitative analyses of specific cyanotoxin accumulation or intoxication in wild birds and mammals in Europe)8,14,25). Commercial kits are also available for determination of cyanotoxins (microcystin) in the tissues and have been used to confirm cyanotoxicosis in birds)12,13).
MANAGEMENT, CONTROL AND REGULATIONS
The temporary management of phycotoxicosis is based on mitigation and/or removal of the bloom, by applying algicides, by physical removal of surface scums and by mixing of the water column. However, possible adverse effects on the ecosystem have to be considered. Exposure to toxic water, by domestic animals for example, should be avoided whenever possible.
Process- based and statistical models have been developed to predict cyanobacterial bloom occurrence, and their concentration and distribution in water bodies can be monitored.
Anthropogenic input of nutrients into sensitive waters largely affects the rate of blooming of harmful algae. Effective long-term control and management of blooms requires reducing eutrophication, including constraints of human- made supply of nutrients, mostly nitrogen and phosphorus. The European Commission, Institute for Environment and Sustainability (IES) Global Environment Monitoring Unit has developed two eutrophication indices, which allow the assessment of the coastal and marine ecosystem’s status and physical sensitivity to eutrophication, as part of the Research Action 2121 (Monitoring and Assessment of Marine Ecosystems))26).
PUBLIC HEALTH CONCERN
Mass populations of toxic cyanobacteria present important environmental and human health hazards. Phycotoxicoses in humans are mainly associated with consumption of contaminated shellfish and include, among others, paralytic shellfish poisoning (caused by a saxitoxin), neurotoxic shellfish poisoning (caused by brevetoxin), and amnesic shellfish poisoning (caused by domic acid). Algal dermatotoxins and gastrointestinal toxins also affect humans.
Hepatotoxins, such as microcystin, cause major liver damage in humans. Nodularin is a carcinogen. Cases may occur simultaneously in humans and animals when both are exposed to the same source.
SIGNIFICANCE AND IMPLICATIONS
FOR ANIMAL HEALTH
Phycotoxicosis may cause important mortality events in wild animal populations, in particular in wild waterfowl.
REFERENCES
1. Adams, J.A., Seaton, D.D., Buchanan, J.B. & Longbottom, M.R. Biological observations associated with the toxic phytoplankton bloom off the east coast. Nature. 1968;220:24-5.
2. Sivonen, K. & Jones, G. Cyanobacterial toxins. In: Bartram, I.C.J. (ed.). Toxic Cyanobacteria In Water — A Guide to their Public Health Consequences, Monitoring and Management. London: WHO, Spon Press; 1999; pp. 56-125.
3. Codd, G.A., Lindsay, J., Young, F.M., Morrison, L.F. & Metcalf, J.S. Harmful cyanobacteria: from mass mortalities to management measures. In: Huisman, J., Matthijs, H.C.P. & Vissler, P.M. (eds). Harmful Cyanobacteria. Dordrecht, Netherlands: Springer; 2005; pp. 1-23.
4. Wobeser, G.A. Diseases of Wild Waterfowl. New York: Plenum Press; 1981.
5. Landsberg, J.H., Vargo, G.A., Flewelling L.J. & Wiley, FE. Algal biotoxins. In: Thomas, N.J., Hunter, D.B. & Atkinson, C.T. (eds). Infectious Diseases of Wild Birds. Ames, Iowa: Blackwell Publishing; 2007; pp. 431-55.
6. Mur, L.R., Skulberg, O.M. & Utkilen, H. Cyanobacteria in the environment. In: Bartram, I.C.J. (ed.). Toxic Cyanobacteria in Water — A Guide to their Public Health Consequences, Monitoring and Management. London: WHO, Spon Press; 1999; pp. 22-5.
7. Stewart, I., Seawright, A.A. & Shaw, G.R. Cyanobacterial poisoning in livestock, wild mammals and birds - an overview. In: Hudnell, H.K. (ed.). Cyanobacterial Harmful Algal Blooms — State of the Science and Research Needs. NewYork: Springer; 2008; pp. 611-37.
8. Handeland, K. & 0stensvik, 0. Microcystin poisoning in roe deer (Capreolus capreolus). Toxicon. 2010;56:1076-8.
9. Henriksen, P., Carmichael, WW, An, J. & Moestrup, 0. Detection of an anatoxin-a(s)-like anticholinesterase in natural blooms and cultures of cyanobacteria/blue-green algae from Danish lakes and in the stomach contents of poisoned birds. Toxicon. 1997;35:901-13.
10. Onodera, H., Oshima, Y., Henriksen, P. &Yasumoto, T. Confirmation of anatoxin- a(s), in the cyanobacterium Anabaena Iemmermannii, as cause of bird kills in Danish lakes. Toxicon. 1997;35:1645-8.
11. Wirsing, B., Hoffmann L. & Heinze, R. et al. First report on the identification of microcystin in a water bloom collected in Belgium. Systematic and Applied Microbiology. 1998;21:23-7.
12. Alonso-Andicoberry, C., Garcia-Villada, L., Lopez-Rodas,V & Costas, E. Catastrophic mortality of flamingos in a Spanish national park caused by cyanobacteria. Veterinary Record. 2002;151:706-7.
13. Lopez-Rodas, V., Maneiro, E., Lanzarot, M.P., Perdigones, N. & Costas, E. Mass wildlife mortality due to cyanobacteria in the Donana National Park, Spain. Veterinary Record. 2008;162:317-8.
14. Sipia, VO., Karlsson, K.M., Meriluoto, J.A.O. & Kankaanpaa, H.T Eiders (Somateria mollissima) obtain nodularin, a cyanobacterial hepa- totoxin, in the Baltic Sea food web. Environmental Toxicology and Chemistry. 2003;23:1256-60.
15. Flewelling, L.J., Naar, J.P., Abbott, J.P. et al. Brevetoxicosis: red tides and marine mammal mortalities. Nature. 2005;435:755-6.
16. Miller, M.M., Kudela, R.M., Mekebri, A. et al. Evidence for a novel marine harmful algal bloom: cyanotoxin (microcystin) transfer from land to sea otters. PLOS One. 2010;5:pii e12576.
17. Skulberg, O.M., Codd, G.A. & Carmichael, WW Toxic blue-green algal blooms in Europe: a growing problem. Ambio. 1984;13:244-7.
18. Bartram, J., Carmichael, W.W., Chorus, I., Jones, G. & Skulberg, O.M. Introduction. In: Chorus, I. & Bartram, J. (eds). Toxic Cyanobacteria in Water — A Guide to their Public Health Consequences, Monitoring and Management. London: WHO, Spon Press; 1999; pp. 12-24.
19. Radostits, O.M., Gay, C.C., Hinchcliff, K.W & Contable, PD. Veterinary Medicine. A Textbook of the Diseases of Cattle, Horses, Sheep, Pigs and Goatt, 10th edn. London: WB Saunders; 2007.
20. Lopez Rodas, V & Costas, E. Preference of mice to consume Microcystis aeruginosa (toxin-producing cyanobacteria): a possible explanation for numerous fatalities of livestock and wildlife. Research in Veterinary Science. 1999;67:107-10.
21. Gunn, G.J., Rafferty, A.G., Rafferty, G.C. et al. Fatal canine neurotoxicosis attributed to blue-green algae (cyanobacteria). Veterinary Record. 1992;130:301-2.
22. Mez, K., Beattie, K.A., Codd, G.A. et al. Identification of a microcystin in benthic cyanobacteria linked to cattle deaths on alpine pastures in Switzerland. European Journal of Phycology. 1997;32:111-7.
23. Falconer, I.R., Buckley, T & Runnegar, M.T.C. Biological half-life, organ distribution and excretion of I- 125 labeled toxic peptide from the blue-green alga Microcystis aeruginosa. Australian Journal of Biological Science. 1986;39:17-21.
24. Hooser, S.B., Beasley, V.R., Waite, L.L., Kuhlenschmidt, M.S., Carmichael, WW & Haschek, WM. Actin filament alterations in rat hepatocytes induced in vivo and in vitro by microcystin-LR, a hepato- toxin from the blue-green alga, Microcystis aeruginosa. Veterinary Pathology. 1991;28:259-66.
25. Karlsson, K.M., Spoof, L.E. & Meriluoto, J.A.O. Quantitative LC- ESI-MS analyses of microcystins and nodularin-R in animal tissue - matrix effects and method validation. Environmental Toxicology. 2005;20:1617-27.
26. Institute for Environment and Sustainability (IES) GEMU. Available online at: http://bioval.jrc.ec.europa.eu [accessed 26 March 2012].
More on the topic CHAPTER 41 HARMFUL ALGAL BLOOMS INCLUDING Cyanobacterial toxicosis:
- Expelling and Avoiding Harmful Spirits
- Harmful Effects of Soil Pollutants
- Tropospheric ozone is harmful to organisms
- Loss of stratospheric ozone increases transmission of harmful radiation
- Reasons for including resource dynamics
- “I shall never, under any circumstances, agree to a representative form of government because I consider it harmful to the people whom God has entrusted to my care.” —Tsar Nicholas II (Alpha History, 2022)
- The optimal transport of each neonatal and pediatric patient is facilitated when the personnel involved ensure the appropriate use of available resources, including staff, equipment, and vehicles.
- The optimal transport of each neonatal and pediatric patient is facilitated when the personnel involved ensure the appropriate use of available resources, including staff, equipment, and vehicles.
- Throughout this book, we have emphasized the role that climate plays in ecological processes, including the distributions and physiological performance of organisms, the rates of resource supply, and the outcomes of biological interactions such as competition.
- Across the country, hospitals and other medical institutions are acknowledging the importanceof including the family in critical medical and mental health decisions.
- “It remains to be seen how soon the reformers will realize the account that must sooner or later be settled between real civil and religious liberty and Mohammedan sacred law or ‘Shariat’ [Sharia] (including the Koran, and the Traditions)....
- The outcome of competition between species can be changed by a broad suite of factors, including features of the physical environment, disturbance, and interactions with other species.
- Pre-Columbian Maya (pre-1502) ritual practices encompassed a range of violent acts generally glossed by the catch-all term ‘sacrifice', including bloodletting and other forms of self-inflicted injury,
- Electromyography (EMG), nerve conduction studies (NCS), and evoked potentials, including somatosensory- evoked potentials (SSEPs)
- Part IV of this handbook applies theories and accounts of collective responsibility to real-world problems, including collective responsibility for: war, global poverty, climate change, conspiracy theories, environmental injustice, and institutional racism.
- The chapters in Part I discuss some of the central debates and theories in the area of collective responsibility including whether collective responsibility should be understood distributively, as attributions of responsibility to group members or non-distributively,
- Financial statement analysis is an essential skill in a variety of occupations including investment management, corporate finance, commercial lending, and the extension of credit. For individuals engaged in such activities, or who analyze financial data in connection with their personal investment decisions, there are two distinct approaches to the task.
- Kidneys are not only the primary excretory organs, but also contribute to many other functions including electrolyte balance, blood pressure regulation, vitamin D metabolism, and secretion of hormones, e.g. erythropoietin and prostaglandins. Apart from primary renal diseases, kidneys are also involved in many systemic disorders or just by being the excretory outlet of body.
- Chapter 2 Refraining from Seeking Clarification: A Chapter from al-Wafl fl sharh al-Wafiya of al-Acraji (d. 1227/1812)
- Chapter 8 Why Early Muslims Divided into Sects? A Chapter from the Mukhtasar al-usul of cAli b. Muhammad b. al-Walid (d. 612/1215)1