TICK-BORNE ENCEPHALITIS
Herbert Weissenb ock
Pathology and Forensic Veterinary Medicine, Department of Pathobiology, University of Veterinary Medicine, Vienna, Austria
Tick-borne encephalitis is a flaviviral nervous system disease of humans, which sporadically also affects several animal species and for which wild mammals serve as reservoir hosts.
AETIOLOGY
Tick - borne encephalitis virus (TBEV) belongs to the family Flaviviridae, genus Flavivirus. The species of TBEV is grouped into three subtypes: the European, the Far Eastern and the Siberian subtypes1-26’27). This assignment is based on their shared pathogenicity for humans rather than on genomic similarities. A different classification ofTBEV has been proposed according to genome sequence data, which includes the assignment of tick- borne encephalitis and louping-ill viruses to the same species (TBEV), of which there are four viral types (Louping-ill virus, including Spanish, British and Irish subtypes; Western tick-borne encephalitis virus (W-TBEV); Eastern tick-borne encephalitis virus (E-TBEV) including far eastern and Siberian subtypes; and Turkish sheep encephalitis virus, including the Greek goat encephalitis virus subtype)(28). Louping-ill has already been described in this chapter.
EPIDEMIOLOGY
The distribution range of W-TBEV is from the Urals in Russia to the Alsace region in France, Scandinavia in the north and parts of the Mediterranean areas along the Adriatic coast to the south. The far eastern subtype is distributed from the Baltic countries to northeastern China and northern Japan, and the Siberian subtype is found in Siberia and the Baltic states. Thus, in parts of the Baltic states the three virus types occur simultaneously. TBEV is not a primary cause of clinical disease in wildlife, but it has a considerable pathogenic potential for humans.
Wild mammals are the most important reservoir hosts and also useful sentinels for estimation of virus activity.For efficient transmission, the virus must be capable of multiplication within the vector. The vector then carries the pathogen to a range of hosts that play different roles within natural foci. The principal vector of W-TBEV is the tick Ixodes ricinus, and there is an overlap in the distribution pattern of this tick species and W-TBEV. In the European part of Russia and in Asia, I. persulcatus is the most prevalent vector, which is correlated with the presence of the E-TBEV strains.
Reservoir hosts of TBEV are small mammals, such as yellow-necked mice (Apodemus flavicollis) and bank voles (Clethrionomys glareolus). Infected ticks, by the mechanism of trans- stadial transmission, remain virus carriers for their entire life and are the most important source of virus. The most relevant transmission event between ticks seems to be co-feeding of several ticks of different developmental stages on one reservoir host. By this mechanism infected tick nymphs are able to transmit the virus to non- infected larvae, which take their blood meal on the same host. Viraemia of the host is not a prerequisite, because the infection occurs by means of infected skin cells. The small mammal hosts have evolution-based adaptation to the virus and do not develop disease. Indicator hosts of TBEV are humans and wild animals larger than rodents, such as hares (Lepus europaeus), roe deer ( Capreo- lus capreolus), foxes (Vulpes vulpes) and wild boar (Sus scrofa). These species become infected accidentally, and although they are not able to transmit the virus back to feeding ticks, they support virus circulation by enabling the ticks themselves to survive and reproduce. These species become infected with TBEV and show seroconversion; thus in endemic areas seroprevalence in large wild mammals can be used as indicators for TBEV transmission within a geographical region. Wild birds are accidental hosts of TBEV.
Birds can become infected and develop viraemia, but generally neither participate in virus circulation nor form a significant nutrition source for ticks(29). Wild birds, according to seroprevalence studies, are infrequently infected with TBEV and have not been shown to play an active role in maintenance of viral activity in endemic foci.Transovarial transmission in ticks has been demonstrated experimentally. This, as well as the vertical transmission by small mammals to their offspring, does not seem to play a major epidemiological role in the transmission of TBEV. TBEV is mainly transmitted by infected ticks, but transmission by unpasteurized milk or milk products from clinically inapparently infected sheep and goats has been repeatedly described.
Seroprevalence between 14% and almost 50% in small mammals and of 10% in large mammals has been shown in different endemic areas in Austria and Germany. In an endemic area in Russia, viral nucleic acid was recovered by RT-PCR in 58% of investigated Sorex araneus shrews, 44% of Clethrionomus rutilus and 20% of Apodemus agrarius mice33'.
PATHOGENESIS, PATHOLOGY AND IMMUNITY
Experimentally, the pathogenesis of TBE is characterized by initial viral replication in endothelial cells and macrophages at the site of the tick bite, followed by transport of the virus to the lymphatic system and viraemia. At this stage, the virus attains access to the nervous system, where it replicates in neurons, leading to their necrosis, and initiates an inflammatory response.
Little information is available on the pathology of natural TBE in wild animals.
A euthanized mouflon ( Ovis ammon musimon) found moribund in a hunting reserve in eastern Austria had a severe infestation with I. ricinus and neuropathological examination revealed a marked non-suppurative lep- tomeningoencephalitis, with the most severe inflammatory changes in the brainstem. Characteristic histological features were neuronal necroses with accompanying neu- ronophagic areas, diffuse microglial activation and numerous lympho-histiocytic perivascular cuffs (Figure 9.6).
IHC demonstrated TBEV antigen in numerous neurons (Figure 9.7) and also in the cytoplasm of macrophages. Partial nucleotide sequencing showed a close relationship to the TBEV strain, Neudorfl, which had been isolated several years previously in the same area(33).A Barbary macaque (Macaca sylvanus) from a group of about 200 animals in a large, outdoor enclosure of a monkey park in a TBE-endemic area in southern Germany
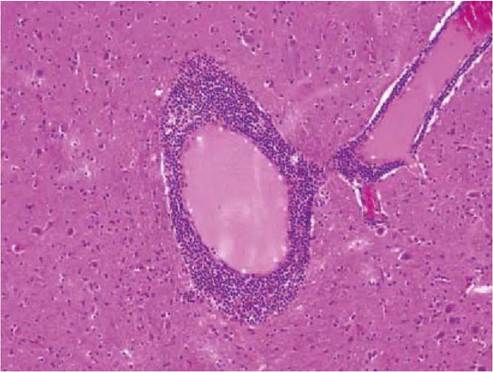
FIGURE 9.6 Severe non-suppurative encephalitis characterized by perivascular cuffs consisting of lymphocytes and monocytes. TBE, mouflon, haematoxylin and eosin.
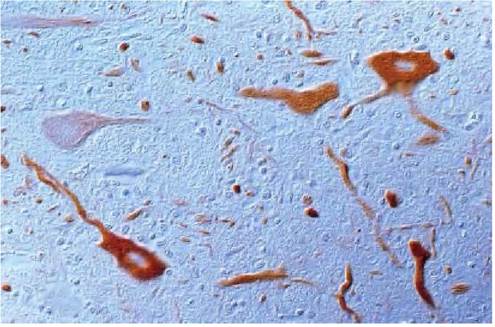
FIGURE 9.7 TBE viral antigen can be demonstrated as brown reaction products in the cytoplasm of neurons and in neuronal processes. Mouflon, IHC.
showed staggering paresis of the hind legs, incoordination and intermittent opisthotonos. Four days after the onset of clinical signs, the animal became comatose and was euthanized. Histologic examination of the brain showed moderate perivascular inflammatory cuffs and mild diffuse infiltration of brain parenchyma with mononuclear cells in almost all brain areas, including basal ganglia and cerebellum. In addition, mild mononuclear inflammatory infiltrates were present in the meninges. Single neuronophagias were also observed. Microglial nodules were not detected. IHC demonstrated viral antigen in several neurons and their processes, including Purkinje cells of the cerebellar cortex and pyramidal neurons of the temporal cortex. Genetic analysis of the entire genome of the isolated TBEV strain showed a typical member ofthe W-TBEV subtype (34).
There are no reports of clinical disease and pathological changes in wild rodents. Experimentally infected laboratory mice show extensive neuronal necroses and encephalitis.
CLINICAL SIGNS
Clinical disease and death due to TBE have been recorded infrequently in domestic animals (dogs, horses, goat, sheep).
Especially in dogs, which next to humans are most susceptible to disease, the clinical signs and the neuropathology are well studied(35). In fatal as well as surviving cases, there is fever and severe neurological signs, such as tremor and ataxia leading to convulsions and epileptiform seizures.DIAGNOSIS
Diagnostic tests for TBE are comparatively straightforward in dead animals but more challenging in clinical samples from living animals. In dead animals, there are neuropatho- logical findings consistent with an infection with a neuronotropic virus, accompanied by the presence of antigen in neurons. In addition, TBEV can be isolated or amplified by RT-PCR from samples of the nervous system. Confirmation of TBE in animals with transient or no clinical signs depends on the detection of viral genome and, more importantly, of specific antibodies. Viral isolation or RT- PCR from either blood or cerebrospinal fluid (CSF) plays a minor role in TBE diagnostics, because the virus is usually cleared from the blood before the onset of CNS clinical signs. Thus, diagnostic assays are mainly based on the detection of specific antibodies. As a rule, TBEV immunoglobulin M (IgM) and usually TBEV IgG antibodies are present in the first serum samples taken when CNS signs become manifest. Intrathecal IgM and IgG antibody response can be detected in CSF, but several days later than in serum. Enzyme immunoassays are usually used for specific serodiagnosis. Also haemagglutination inhibition is widely used, but it measures all antibody classes and requires a rise in antibody titre for definitive diagnosis. Because of high cross-reactivity of the antigenic structure in the flavivirus, possible diagnostic difficulties could arise in areas where other flaviviruses co-circulate (e.g. West Nile virus). In such cases, neutralization tests for the confirmation ofTBEV specificity of the antibodies are needed(36).
MANAGEMENT, CONTROL
AND REGULATIONS
The vaccines against TBE are only licensed for use in humans and not for animals.
As the virus is maintained in transmission cycles between small rodents and ticks, once infected a tick carries the virus for the rest of its life; therefore the elimination of the virus from natural endemic foci is not feasible.PUBLIC HEALTH CONCERN
The risk of acquiring TBE is correlated with the likelihood of tick exposure; thus the infection rate in endemic areas is higher in those individuals who have more frequent outdoor activities. The infection rate within the population of a country in which TBE is endemic can be markedly reduced by vaccination programmes. This has been shown in Austria, where a large percentage of inhabitants are vaccinated against TBE.
Apart from Japanese encephalitis virus, TBEV is the encephalitogenic flavivirus with the most severe impact on human health in Europe and Asia. Each year it is responsible for several thousands cases of encephalitis in humans, and the fatality rate is significantly higher with the eastern subtypes (up to 20%), than with the western subtype (approximately 1%).
More on the topic TICK-BORNE ENCEPHALITIS:
- 2 SELECTED ZOONOTIC PATHOGENS WITH EUROPEAN WILDLIFE RESERVOIRS/HOSTS
- THE MITES AND TICKS
- Mosquito-Borne Illnesses: Western Nile Virus
- 5 Appendices
- Pregnant Women and Chemical-Biological Warfare