Optimal foraging theory addresses behavioral choices that enhance the rate of energy gain
Animals face several challenges when searching for food (Concept 5.4). The abundance of food and its nutritional content vary in space and time. Additionally, how easy the food items are to detect, capture, or subdue can impact an animal's success.
These factors will influence the nutritional benefit and the amount of energy that must be expended to acquire food, and therefore its net benefit to the animal. For example, a lizard foraging for ants may find higher abundances of ants near the colony's tunnels and an abrupt decrease when moving away from the ant domicile. Ant species also vary in their concentrations of nitrogen and carbon and in their abilities to defend against predators. Feeding in areas with the most dense assembly of ant species with the highest nutrient concentrations and the greatest ease of capture would provide the greatest benefit to the foraging lizard.Ecologists have theorized that foraging efficiency can be viewed as the energy benefit per amount of energy expended to find food. An optimal solution to the challenges posed above would result in the maximum energy gained per unit of energy expended. If energy is in short supply and impacts an animal's fitness, then selection should act on foraging behavior to increase efficiency. Optimal foraging theory was developed by several ecologists in the 1960s and 1970s as a framework to develop testable hypotheses of foraging behavior (MacArthur and Pianka 1966; Pyke 1984). This approach has been used for several aspects of foraging, focusing primarily on food patchiness, quality, and the ease of acquisition.
Optimal foraging theory provides an opportunity to use straightforward mathematical models to evaluate hypotheses associated with adaptive behavior. In simple mathematical terms, optimal foraging can be represented by the following equation:
Enet = Egross/ (h + s)
where Enet is the net energy obtained from a food source, Egross is the total energy from a food source, h is the time spent handling the food source, and s is the time spent searching for the food.
The net energy value is determined by the energy value of the food item as well as the energy expended in handling and searching for it. Food that requires substantial handling (e.g., a tough nut to crack or a fighting prey) will yield lower net energy than food that requires less handling. Likewise, food that is sparsely distributed will require greater searching time than food that is more abundant.Simple conceptual models that describe the net amount of energy that an animal gets from its food can also be used to formulate predictions of optimal foraging (FIGURE 8.6). As an animal increases its effort to forage (the time and energy it spends searching for, capturing, subduing, and consuming food), the total amount of energy obtained (blue curve) initially increases rapidly. At some point, however, a further increase in foraging effort provides relatively little additional energy, and the net energy gain begins to decrease. Several factors may cause this decrease, including a limitation on how much food the animal can carry or ingest.
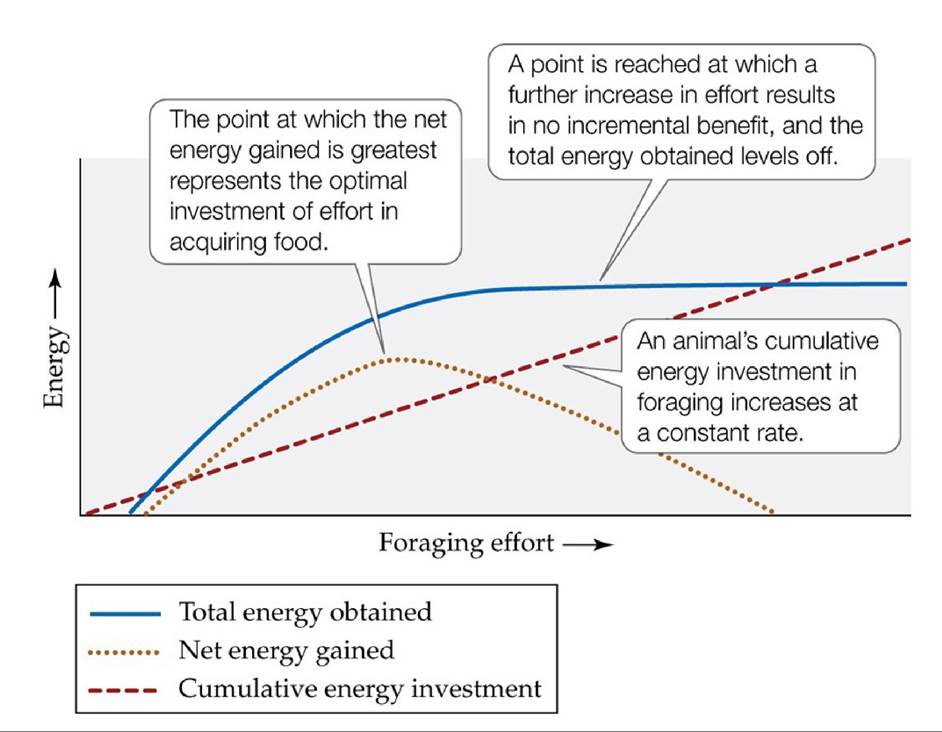
FIGURE 8.6 Conceptual Model of Optimal Foraging The net energy gained from foraging (dotted curve) equals the total energy obtained from the food acquired (solid curve) minus the cumulative energy invested in acquiring that food (dashed curve). This simple model can be used to test whether animals forage in a manner that results in the maximum benefit, based on estimates for the total energy obtained and the cumulative energy invested.
Suppose you could estimate the net energy gained at different levels of foraging effort expended by lizards eating ants in the desert. How could you use that information to test whether the lizards foraged optimally?
(After G. Parker and J. Smith. 1990. Nature 348: 27-33.) View larger image
While the models discussed here are simple, they provide a basis for making quantitative predictions about animal foraging behavior.
More sophisticated models have been used to derive hypotheses that can be tested under field or laboratory conditions. An important component of these models is the currency (such as net energy gain) that is used to determine the benefit. Such models might incorporate, for example, net energy gained, time spent feeding, and risk of predation (Schoener 1971). If foraging behavior is an adaptation to limited food supplies, then we must be able to relate the benefit of that behavior to the survival and reproduction of the animal.Tests of Optimal Foraging Theory
Research addressing optimal foraging has focused on diet selection, selection of patches to feed in, time spent in food patches, and prey movements (Pyke 2019). In one of the key early tests, John Krebs et al. (1977) devised a unique way to evaluate whether great tits (Parus major), a common bird found throughout much of Eurasia and northern Africa, selected prey types of greatest profitability (energy gained per unit of handling time). They placed captive birds next to a moving conveyer belt carrying prey that differed in size (large and small mealworms) and in the time required to obtain them (each of the small mealworms was taped to the surface of the conveyer belt, thus requiring greater handling time, which decreased their profitability). By changing the proportions of the prey types and the distances between adjacent prey on the conveyer belt (variation in effort associated with search time), the researchers varied the profitability of the large and small mealworms. Using a model of optimal foraging and measurements of the times it took individual birds to subdue and consume the prey (handling time), they predicted how frequently the birds should select the large mealworms as encounter rates with the two prey types were varied. The birds consumed an increasing percentage of large mealworms as the relative profitability of those larger prey increased (FIGURE 8.7), just as the model predicted.
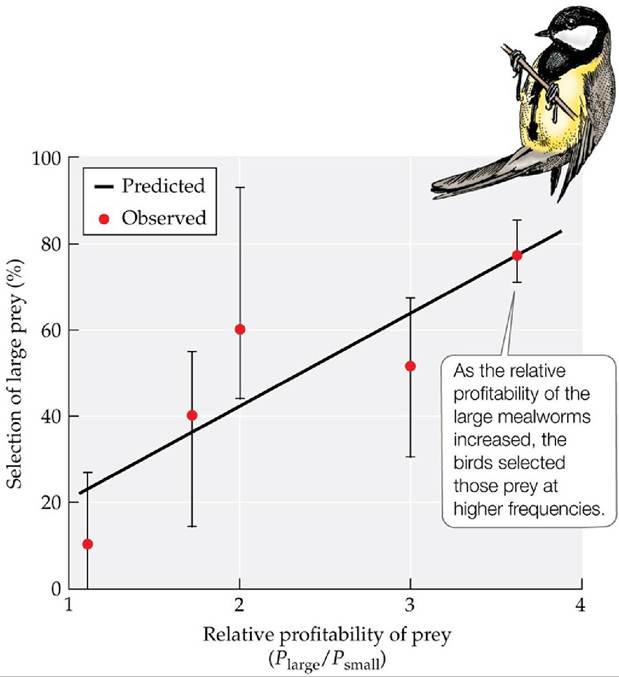
FIGURE 8.7 Effect of Profitability on Food Selection Krebs et al.
used an optimal diet selection model, along with measurements of prey handling time for individual birds, to predict the rate at which great tits (Parus major) would select large over small mealworms as their encounter rates with the two prey types were varied (expressed as the calculated ratio of profitabilities of the prey types), where profitability refers to the energy per unit of handling time. Error bars show ± one SE of the mean. (After J. B. Krebs et al. 1977. Anim Behav 25: 30-38.) View larger imageA field study by Meire and Ervynck (1986) focused on the diet selection of the Eurasian oystercatcher (Haematopus ostralegus), a shorebird that eats bivalves (e.g., clams and mussels). Oystercatchers must find a bivalve buried in the sand, lift it out, and open it before they can eat it. For bivalves below a certain size, the net energy gain from this effort is small, setting a lower limit on the bivalve size selected by the oystercatchers. Bivalves above a certain size have thicker shells and require more effort to open, setting an upper limit on the bivalve size selected by the birds. Meire and Ervynck demonstrated that oystercatchers select prey of sizes that fall between these limits, which provide the most energy gain for the effort, despite the relatively low abundance of prey of these sizes.
The Marginal Value Theorem
Another aspect of optimal foraging theory considers the habitat in which an animal forages as a heterogeneous landscape made up of patches containing different amounts of food. To optimize its energy gain, an animal should forage in the most profitable patches—those in which it can achieve the highest energy gain per unit of time. We can also consider the benefit obtained by a foraging animal from the perspective of time spent in a patch. Once the forager finds a profitable patch, its rate of energy gain is initially high, but that rate decreases and eventually becomes marginal as the forager depletes the food supply. Simple conceptual models predict that a foraging animal should stay in a patch until the time when the rate of energy gain in that patch has declined to the average rate for the habitat (known as the giving-up time), then depart for another patch.
The giving-up time should also be influenced by the distance to other patches. Effort must be invested in traveling to another patch, so the animal may accept a lower rate of energy gain if the distance between patches is greater. This conceptual model, called the marginal value theorem, was initially developed by Eric Charnov (1976). It can be used to evaluate the influences of distance between patches, the quality of the food in a patch, and the animal's energy-extraction efficiency on the giving-up time. The model has also been extended to other “giving-up” problems in behavioral ecology, including how long to copulate and when to cease guarding a nest and seek other mates.One of the predictions of the marginal value theorem is that the longer the travel time between food patches, the longer an animal should spend in a patch. This prediction was tested by Richard Cowie (1977) using a laboratory setup with great tits in a “forest” composed of wooden dowels. The food “patches” consisted of sawdust-filled plastic cups containing mealworms. The “travel time” among patches was manipulated by placing cardboard covers on top of the food cups and adjusting the ease with which they could be removed by the birds. Cowie used the marginal value theorem to predict the amount of time the birds should spend in the patches, based on the travel time between them. His results matched the model predictions fairly well (Cowie 1977) (FIGURE 8.8). Similar results have been obtained from other laboratory experiments as well as from studies using observational studies of foragers, such as Munger's 1984 study on the behavior of horned lizards (Phrynosoma spp.) foraging for ants in the Chihuahuan Desert.
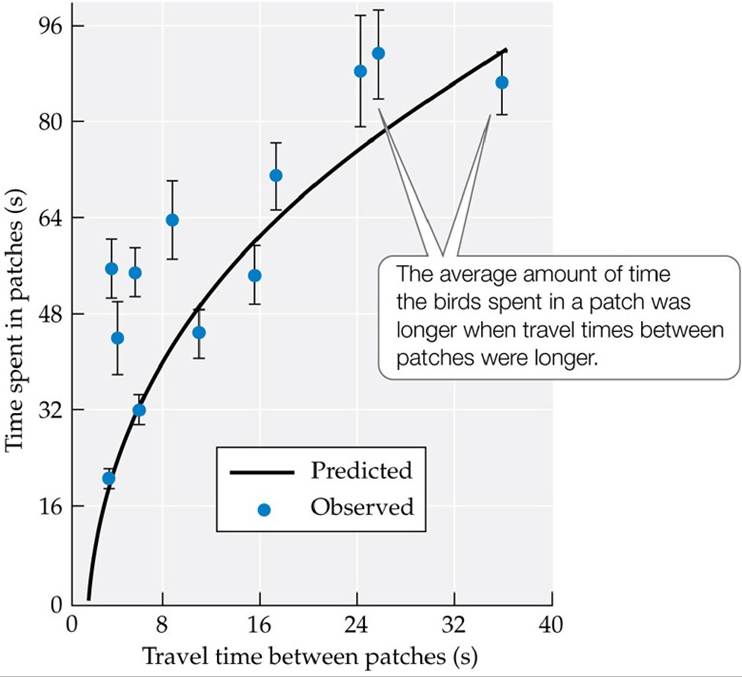
FIGURE 8.8 Effect of Travel Time between Patches Inalaboratoryexperiment1Cowie used the marginal value theorem to predict how the travel time between patches would affect the average amount of time great tits (Parus major) spent in a patch.
Error bars show ± one SE of themean. (After R. J. Cowie. 1977. Nature 268: 137-139.) View larger image
The marginal value theorem also predicts that the time spent in food patches is positively related to their quality. Watanabe et al. (2014) studied foraging rates of Adelie penguins (Pygoscelis adeliae) on krill (Euphausia Crystallorophias) in Antarctic waters. Using animal-borne video cameras and sensors to detect rapid head movement associated with capturing krill, they tested whether foraging penguins spent more time in denser patches of krill. Their results supported the marginal value theorem's prediction that in the short term (individual dives), the penguins spent more time in patches with higher krill abundance (FIGURE 8.9). This and other studies using methods that remotely track foraging animals and detect prey capture have led to model refinements that incorporate energetic costs of capturing prey in the consideration of time spent in patches (Foo et al. 2016).
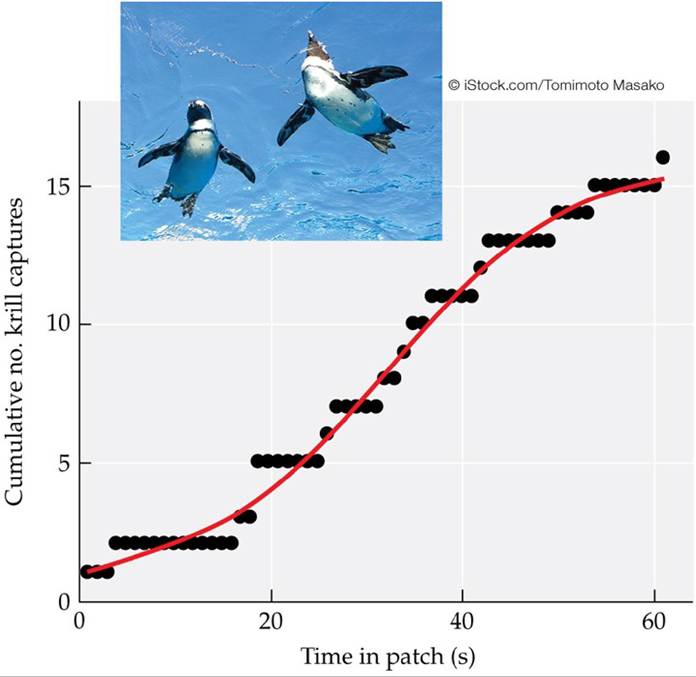
FIGURE 8.9 ForaginginAdeliePenguins The relationship between the amount of time spent in a patch of krill (Euphausia Crystallorophias) by foraging Adelie penguins (Pygoscelis adeliae) versus the richness of the patch (as estimated by the rate of krill captures). The data support the prediction of the marginal value theorem that foragers will spend greater time in richer patches than in those with fewer prey. (After Y. Y. Watanabe et al. 2014. Proc R Soc B 281: 20132376.) View larger image
Research testing optimal foraging theory has increasingly incorporated additional complexities into its models and predictions, including risk of predation and environmental stress, foraging experience of the animal, and interactions with other species (competition, predation, facilitation). Generally, optimal foraging theory has been successful at supporting qualitative predictions (descriptions of behavior construed as consistent with the theory, such as eating a more diverse diet under certain conditions) but less so with more quantitative ones (predictions based on measurements of foraging behavior, such as the average time spent feeding in a patch) (Pyke 2019). Optimal foraging theory best describes the foraging behavior of animals that feed on immobile prey and applies less well to animals feeding on mobile prey (Sih and Christensen 2001). In addition, the assumptions that energy is always in short supply and that a shortage of energy dictates foraging behavior may not always be correct. Carnivores, in particular, may not lack for food resources to the degree assumed in optimal foraging models (Jeschke 2007). Furthermore, resources other than energy may be involved in the selection of food items, particularly nutrients such as nitrogen and sodium. And as we'll see next, additional considerations for foragers include the risk of predation and the defenses of prey.
More on the topic Optimal foraging theory addresses behavioral choices that enhance the rate of energy gain:
- CONCEPT 8.2 Animals make behavioral choices that enhance their energy gain and reduce their risk of becoming prey.
- The Optimal Inflation Rate
- Anything that influences energy gain by photosynthesis has the potential to affect the survival, growth, and reproduction of the organism.
- The second law of thermodynamics states that during any transfer of energy, some energy is dispersed as unusable energy because of the tendency toward an increase in disorder (entropy).
- Chapter 16 Behavioral Finance in Theory and Practice
- Chapter 3 An Agent-Based Model of Resource Distribution on Hunter-Gatherer Foraging Strategies: Clumped Habitats Favor Lower Mobility, but Result in Higher Foraging Returns
- BEHAVIORAL FINANCE: CONTRIBUTIONS TO MODERN FINANCE THEORY
- Energy flow between trophic levels can be depicted using energy or biomass pyramids
- The theory of vague propositions outlined in chapter 6 appealed freely to the orthodox Bayesian theory of credences and learning, which assumes, among other things, that the correct theory of rational credence is one governed by the classical axioms of probability theory.
- The choices of Court
- The vast majority of the autotrophic production of chemical energy on Earth occurs through photosynthesis, a process that uses sunlight to provide the energy needed to take up carbon dioxide and synthesize organic compounds, principally carbohydrates.
- Individuals often alter their foraging decisions when predators are present
- CONCEPT 5.2 Radiant and chemical energy captured by autotrophs is converted into stored energy in carbon-carbon bonds.