Photosynthesis is the powerhouse for life on Earth
Prior to 1650, most people believed that plants obtained the raw material needed for their growth from the soil. Jan Baptist van Helmont (1579-1644), a Flemish scientist, tested this theory experimentally.
He carefully measured the mass of dry soil in a pot (200 pounds or 91 kg) and then planted a willow sapling weighing 5 pounds (2.3 kg). Van Helmont watered the sapling using only rainwater for 5 years as it grew into a small tree. At the end of that time, the tree had gained 164 pounds (74 kg), and the soil had lost only 2 ounces (0.06 kg). Although he incorrectly concluded that the tree had gained its mass from the water, van Helmont's experiment established the basis for the later discovery that photosynthetic uptake of CO2 from the air—not material from the soil—was the source of the tree's weight gain.The vast majority of biologically available energy on Earth is derived from the conversion of sunlight into energy-rich carbon compounds by photosynthesis. Photosynthetic organisms include some archaea, bacteria, and protists and most algae and plants. Leaves are the principal photosynthetic tissue in plants, but photosynthesis may also occur in stem and reproductive tissues. Like chemosynthesis, photosynthesis involves the conversion of CO2 into carbohydrates that are used for energy storage and biosynthesis. Photosynthesis is also responsible for the largest movements of CO2 between Earth and the atmosphere, and it is therefore critically important to the global climate system and climate change (as we'll see in Concept 25.1). Here, we will briefly review the major steps of plant photosynthesis and consider some ecologically relevant constraints on photosynthetic rates. In Concept 5.3, we will examine some variations in plant photosynthetic pathways.
Light-Driven and Carbon Reactions
Photosynthesis has two major steps.
The first is the harvesting of energy from sunlight, which is used to split water to provide electrons for generating ATP and NADPH. This step is often referred to as the light-driven reactions of photosynthesis. The second step is the fixation of carbon and the synthesis of sugars and subsequently carbohydrates. This step is often referred to as the carbon reactions of photosynthesis.Sunlight harvesting is accomplished by several pigments, principally chlorophyll. Chlorophyll gives photosynthetic organisms their green appearance because it absorbs red and blue light and reflects green wavelengths (FIGURE 5.6). Plants and photosynthetic bacteria have similar chlorophyll pigments, but they absorb light at slightly different wavelengths. Additional pigments associated with photosynthesis, called accessory pigments, include the carotenoids, which are characteristically red, yellow, or orange in appearance. All of these photosynthetic pigments are embedded in a membrane, along with other molecules involved in the light-driven reactions. In plants, this membrane lies within specialized organelles called chloroplasts, while in photosynthetic bacteria the pigments are embedded in the cell membrane. The pigment molecules are arrayed like antennae, with each array containing 50-300 molecules. The pigments absorb energy from discrete units of light, called photons. That energy is used to split water and provide electrons. The electrons are passed on to molecular complexes on the membranes, where they are used to synthesize ATP and NADPH.
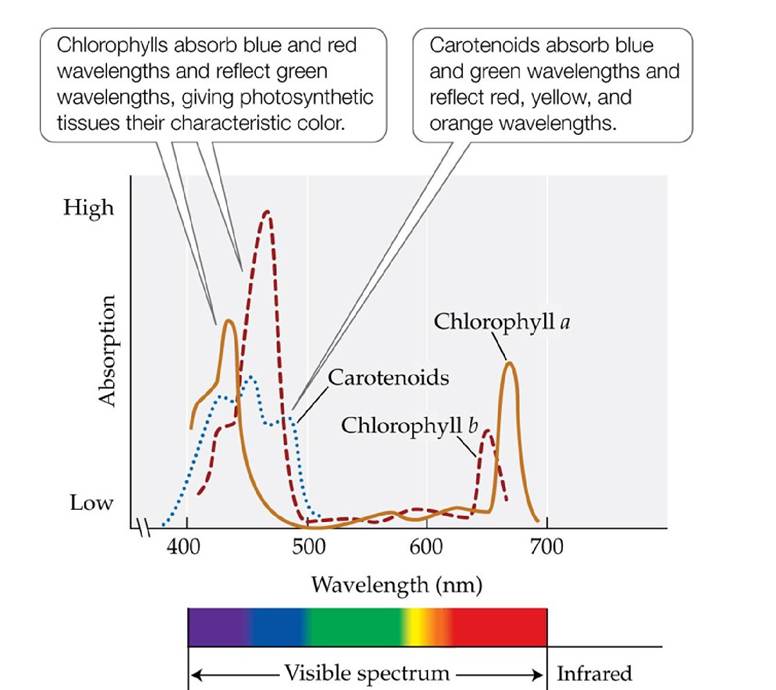
FIGURE 5.6 Absorption Spectra of Plant Photosynthetic Pigments Plantstypically contain several light-absorbing pigments, which absorb light of different wavelengths. (After C. J.
Avers. 1985. Molecular Cell Biology. Addison-Wesley: Boston, MA.) View larger image
The splitting of water (H2O) to provide electrons for the light-driven reactions generates oxygen (O2).
The evolution of photosynthesis, and the accompanying release of O2 into the atmosphere, was a critical step in the development of the chemistry of the modern atmosphere and lithosphere as well as the evolution of life on Earth. Atmospheric oxygen led to the creation of a layer of ozone (O3) high in the atmosphere that shields organisms from high-energy ultraviolet radiation (described in Concept 25.4). The evolution of aerobic respiration, in which O2 is used as an electron acceptor, facilitated great evolutionary changes for life on Earth.In the carbon reactions of photosynthesis, energy from ATP and NADPH is used in the Calvin cycle to fix carbon. Carbon dioxide is taken up from the atmosphere through the stomates of vascular plants, or it diffuses across the cell membranes in nonvascular plants, algae, and photosynthetic bacteria and archaea. A key enzyme associated with the Calvin cycle is ribulose 1,5 bisphosphate carboxylase/oxygenase, thankfully usually referred to by its abbreviation, rubisco. Rubisco, the most abundant enzyme on Earth, catalyzes the uptake of CO2 and the synthesis of a three-carbon compound: phosphoglyceraldehyde, or PGA. PGA is eventually converted into a six-carbon sugar [glucose (C6H12O6) in most plants]. The net reaction of photosynthesis is therefore
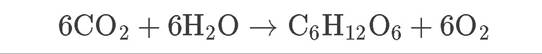
(5.1)
Environmental Constraints and Solutions
The rate of photosynthesis determines the supply of energy and substrates for biosynthesis available in the environment. Because this rate influences the growth and reproduction of photosynthetic organisms—often equated with their ecological success (their abundance and geographic range)—environmental controls on the rate of photosynthesis are a key topic in physiological ecology. It should be noted, however, that net energy (carbon) gain is also influenced by CO2 losses associated with cellular respiration.
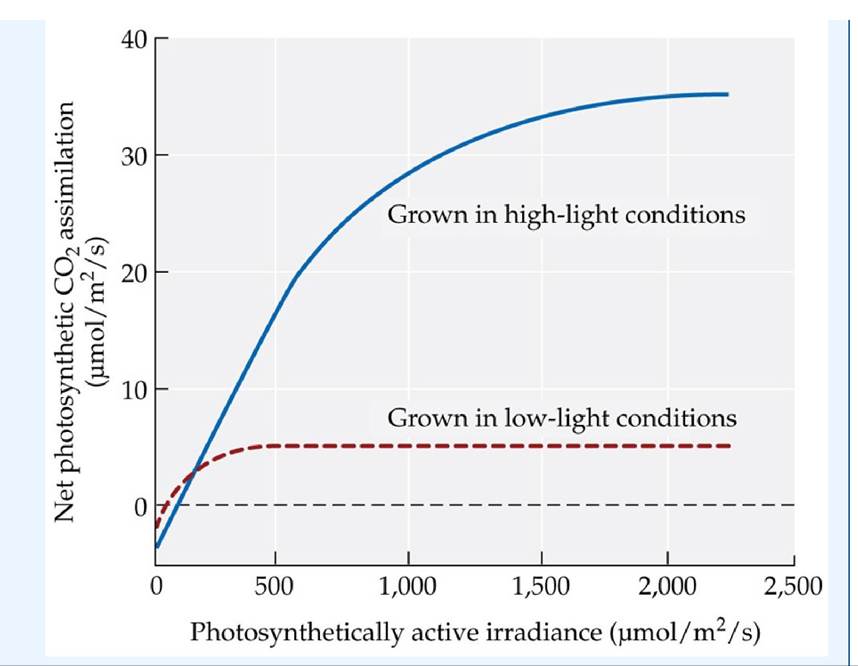
Light is clearly an important influence on rates of photosynthesis in both terrestrial and aquatic habitats. The relationship between the light level and a plant's photosynthetic rate can be portrayed by a light response curve (FIGURE
5.7A). When there is enough light that the plant’s photosynthetic CO2 uptake is balanced by its CO2 loss by respiration, the plant is said to have reached the light compensation point. As the light level increases above the light compensation point, the photosynthetic rate also increases; in other words, photosynthesis is limited by the availability of light. The photosynthetic rate levels off at a light saturation point, which is typically reached at a level below full sunlight.
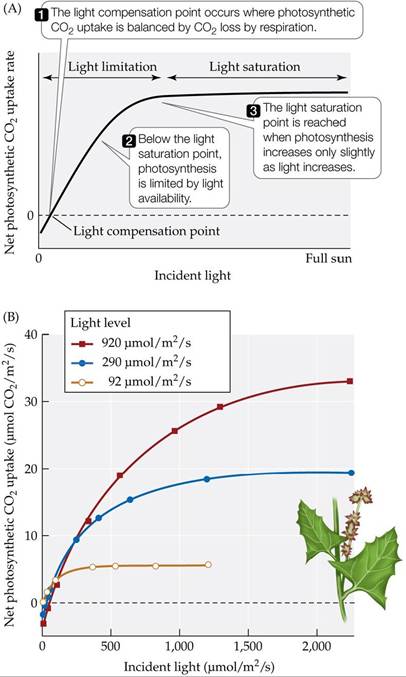
FIGURE 5.7 Plant Responses to Variations in Light Levels (A) Photosynthetic light response curve. (B) Spearscale (Atriplex triangularis) plants grown at different light levels in growth chambers acclimatized to those light levels. Their light response curves indicate that adjustments in the light saturation point occurred. Small, but ecologically significant, changes in the light compensation point occur in many other species, facilitating CO2 uptake at low light levels.
Why might the light saturation point of a plant be below the maximum light level the plant is likely to be exposed to?
(B after O. Bjorkman. 1981. In Physiological Plant Ecology I: Encyclopedia of Plant Physiology, O. L. Lange et al. [Eds.], pp. 57-101. Springer: New York.) View larger image
How do plants cope with light variation? For example, how would an understory forest plant respond to shading by canopy trees? Could that plant acclimatize to more light if the canopy tree fell, allowing full sunlight to reach the ground? In a series of classic studies using controlled growth conditions, Olle Bjorkman demonstrated that acclimatization to different light levels involves a shift in the light saturation point (Bjorkman 1981) (FIGURE 5.7B).
Morphological changes associated with this acclimatization include alterations in the thickness of leaves and variation in the number of chloroplasts available to harvest light (FIGURE 5.8). Photosynthetic organisms may also alter the density of their lightharvesting pigments—a strategy analogous to changing the size of the antenna on a radio—and the amounts of photosynthetic enzymes available for the carbon reactions. Typically, the average light level a plant experiences, integrated over the course of the day, is near the transition point between light limitation and light saturation (see ANALYZING DATA 5.1).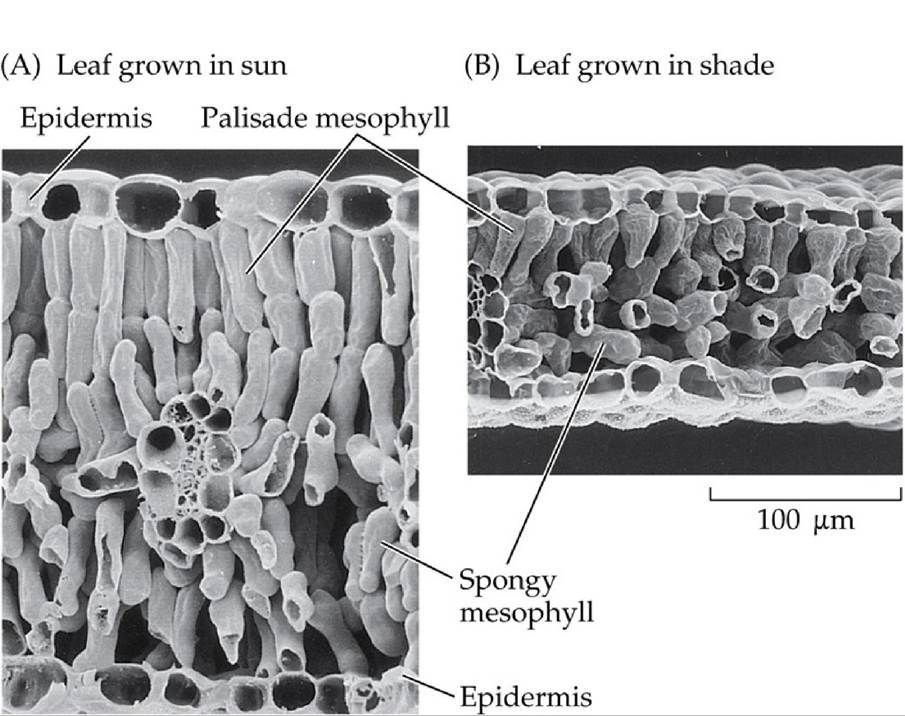
Both photos courtesy of T. Vogelmann
FIGURE 5.8 Effects of Light Level on Leaf Structure Golden banner (Thermopsis montana) leaves adjust morphologically to changes in light levels. Leaves grown at high light levels (A) are thicker, have more photosynthetic cells (palisade and spongy mesophyll), and have greater numbers of chloroplasts than leaves grown at low light levels (B). View larger image
ANALYZING DATA 5.1
How Does Acclimatization Affect Plant Energy Balance?
Many plants can adjust their morphology and biochemistry to match the light conditions under which they are grown. The curves depicted in the figure are from Olle Bjorkman’s* classic studies and show the net photosynthetic CO2 uptake for spearscale plants (Atriplex triangularis) grown under high-light (920 μmol∕m2∕s of photosynthetically active radiation) and low-light (92 μmol∕m2∕s) conditions.
1. Assuming no further physiological changes occur, calculate the daily carbon balance for leaves of the high-light and low- light plants grown under the following conditions:
a. Plants are kept at a light level (irradiance) of 200 μmol∕m2∕s for 2 hours, then switched to an irradiance of 1,500 μmol∕m2∕s for 10 hours, then switched back to 200 μmol∕m2∕s for 2 hours.
The lights are then turned off for 10 hours. (This light regime approximates sunny conditions in an open subtropical savanna.)b. Plants are kept at an irradiance of 50 μmol∕m2∕s for 2 hours, then switched to an irradiance of 200 μmol∕m2∕s for 10 hours, then switched back to 50 μmol∕m2∕s for 2 hours. The lights are then turned off for 10 hours. (This light regime is similar to that expected in a tropical rainforest understory.)
2. High-light and low-light plants exhibit differences in maximum net photosynthesis rates, light compensation points, and nighttime respiration. Which of these three differences contributes the most to the distinction in carbon balance under high-light conditions (calculated in part a of question 1) and low-light conditions (calculated in part b)?
3. What do you think might contribute to the differences in nighttime respiration rates?
*Bjorkman, O. 1981. Responses to different quantum flux densities. In Physiological Plant
Ecology I: Encyclopedia of Plant Physiology, O. L. Lange et al. (Eds.), pp. 57-101. Springer-Verlag, Berlin.
Some specialized bacteria are especially well adapted to photosynthesis at low light levels, which allows them to thrive in dimly lit environments such as relatively deep ocean water (down to about 20 m). A previously undescribed form of chlorophyll, called chlorophyll f, was found in samples of the marine cyanobacteria that form sediments in the shallow waters of Shark Bay, Australia (Chen et al. 2010), and has subsequently been found in cyanobacteria of other low-light habitats, including hot springs, rice paddies, and caves. Chlorophyll f absorbs light in the near-infrared region, just beyond the red wavelengths used by other forms of chlorophyll (see Figure 5.6). Chlorophyll f is an adaptation that allows cyanobacteria possessing it to grow underneath other photosynthetic organisms that use light in the blue and red wavelengths, as it lets them harvest energy at wavelengths that pass through those other photosynthetic organisms (Nurnberg et al. 2018). The discovery of a pigment that can harvest near-infrared energy has implications for increasing the efficiency of photovoltaic panels used to generate electricity, which may help lower emissions of CO2.
Water availability is an important control on the supply of CO2 for photosynthesis in terrestrial plants. As we saw in Concept 4.3, low water availability results in closure of the stomates, restricting the entry of CO2 into leaves. Stomatal control represents an important trade-off for the plant: water conservation versus energy gain through photosynthesis as well as cooling of the leaf through transpiration. Keeping stomates open while tissues lose water can permanently impair physiological processes in the leaf. Closing stomates, however, not only limits photosynthetic CO2 uptake, but also increases the chances of light damage to the leaf. When the Calvin cycle is not operating, energy continues to accumulate in the light-harvesting arrays, and if enough energy builds up, it can damage the photosynthetic membranes. Plants have evolved a number of ways of dissipating this energy safely, including the use of carotenoids to release it as heat.
Temperature influences photosynthesis in two main ways: through its effects on the rates of chemical reactions and by influencing the structural integrity of membranes and enzymes. Autotrophs acclimatize and adapt to temperature variation by changing properties of the Calvin cycle enzymes and/or the photosynthetic membranes. Different photosynthetic organisms have different forms of the same photosynthetic enzymes that operate best under the environmental temperatures where the organisms occur. These differences result in markedly different temperature ranges for photosynthesis in organisms from different climates (FIGURE 5.9A). Lichens and plants of Arctic and alpine environments can photosynthesize at temperatures close to freezing, while desert plants may have their highest photosynthetic rates at temperatures that are hot enough to denature most other plants' enzymes (40oC-50oC or 104°F-122°F). Plants that acclimatize to changes in temperature synthesize different forms of photosynthetic enzymes with different temperature optima (FIGURE 5.9B). Temperature also influences the fluidity of the cell and organelle membranes (see Concept 4.2). Cold sensitivity in plants of tropical and subtropical biomes is associated with loss of membrane fluidity, which inhibits the functioning of the light-harvesting molecules embedded in the chloroplast membranes. And as we have seen, high temperatures, particularly in combination with intense sunlight, can damage photosynthetic membranes.
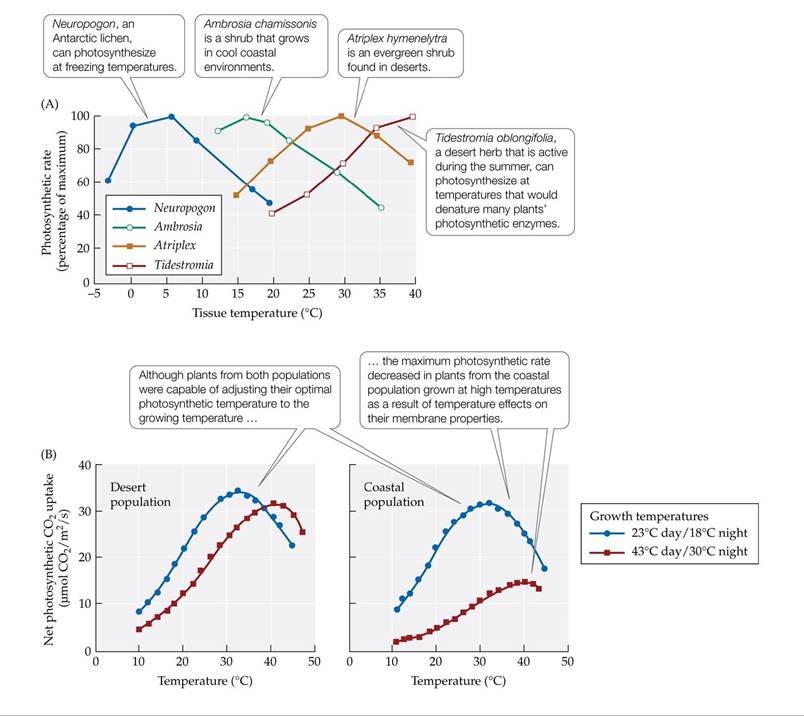
FIGURE 5.9 Photosynthetic Responses to Temperature (A) The temperatures at which plants and lichens reach their maximum photosynthetic rates correspond to the range of environmental temperatures in the native habitat of the species. (B) Acclimatization to different growth temperature regimes by plants from different populations of Atriplex Ientiformis, a shrub that occurs in the hot Mojave Desert and in cool coastal zones of California. The two growth temperature regimes are representative of the two habitats the species occupies. (A after H. A. Mooney. 1986. In Plant Ecology, M. J. Crawley [Ed.]. Blackwell Science Ltd: Oxford. Based on O. L. Lange and L. Kappen. 1972. Antarctic Research Series 20: 80-95. American Geophysical Union; H. A. Mooney et al. 1983. Oecologia 57: 38-42; H. A. Mooney et al. 1976. Carnegie Institution Year Book 75: 410-413. B after R. W. Pearcy. 1977. PlantPhysiol 59: 795-799.) View larger image
Nutrient concentrations in leaves reflect their photosynthetic potential because most of the nitrogen in plants is associated with rubisco and other photosynthetic enzymes. Thus, higher amounts of nitrogen in leaves are correlated with higher photosynthetic rates. Why, then, don't all plants allocate more nitrogen to their leaves to increase their photosynthetic capacity? There are two main reasons. First, the supply of nitrogen is low relative to the demand, and nitrogen is needed for growth and other metabolic functions in addition to photosynthesis (see Chapter 22). Second, increasing the nitrogen concentration of a leaf increases the risk that herbivores will consume the leaf, as plant-eating animals are often nitrogen starved (see Concept 22.1). Plants must balance the competing demands of photosynthesis, growth, and protection from herbivores.
Over evolutionary time, some plants have dealt with environmental constraints on photosynthesis with adaptations in their photosynthetic pathways, as we will see next.
2
- Oxidation-reduction reactions involve the exchange of electrons between chemical compounds. The compound that gives up, or donates, electrons is oxidized, while the compound that accepts electrons is reduced.
More on the topic Photosynthesis is the powerhouse for life on Earth:
- Photosynthesis is the powerhouse for life on Earth
- Photorespiration lowers the efficiency of photosynthesis
- Reviewers