EPIDEMIOLOGY
GEOGRAPHICAL DISTRIBUTION
Until recently, the global distribution of BTV was thought to lie approximately between 35°S and 40°N. Within these bounds it has a worldwide distribution occurring in the Americas, Africa, the Middle East, the Indian subcontinent, China, South-East Asia and northern Australia.
Historically, it also occasionally invaded Iberia and a few Greek islands. Cyprus is the only area of Europe where BTV has occurred regularly.S ubsequent to the 1979—1980 outbreaks in Greece (Lesbos, Rhodes), BTV remained absent from Europe (excluding Cyprus) for almost 20 years. However, in late 1998 new incursions commenced, mainly affecting domestic ruminants. The outbreaks started in several Greek islands close to Turkey but spread to include European Turkey, mainland Greece, Bulgaria, Macedonia, Albania, Bosnia, Croatia, Montenegro and Serbia. There were several unique features to this incursion. First, it involved four serotypes of BTV (1,4, 9 and 16) whereas all previous incursions into Europe had involved only a single serotype at a time. Second, serotypes 1, 9 and 16 had never previously entered Europe; and third, the incursion extended to the River Sava in Serbia, which at 44 ° 50 'N is almost 500 km further north than BTV had ever been recorded in Europe(2).
I n 2000 BTV was also recorded for the first time on Italian territory, in Sardinia. The origin of the outbreak was probably northern Tunisia, where the virus had been active since late 1999(3). This incursion was due to BTV2 and was the first occasion this serotype had been recorded in Europe. It spread from Sardinia to Sicily and to several regions on mainland Italy. In the period 2002—2003 a new phase of the incursion commenced when, in addition to BTV2, BTV4, BTV9 and BTV16 were also identified and disease was detected in eight provinces in Italy(3).
BTV4, BTV9 and BTV16 had already been reported from the eastern Mediterranean Basin (see above) but the route of incursion into Italy is uncertain, although illegal movement of animals is suspected. Outbreaks due to one or more of these serotypes have continued in some Italian provinces until the time of writing (2010).In late 2000, BTV invaded French territory for the first time, when BTV2 was identified in Corsica and the outbreak continued until mid-2001. In 2003, a further incursion into Corsica occurred when BTV4 was identified, and this was followed in 2004 by BTV16.
In September—October 2000 the first BTV incursions into Spanish territory since 1960 commenced. The initial outbreaks, on the Balearic islands, were due to BTV2, although in 2004 BTV4 was also isolated there(4). Subsequent incursions of BTV4, from 2004, involved wide swaths of mainland Spain and Portugal. This latter incursion probably originated in Morocco and/or Algeria and after starting in southern mainland Spain spread northwards and westwards. In 2007 a fresh BTV incursion from North Africa occurred. This involved BTV1, and the virus spread northwards, reaching South West France in late
2007 and the north coast of France in late 2008.
Separate from the BTV incursions into southern and eastern Europe, and the incremental progression northwards of some serotypes, in 2006, an incursion of BTV8 directly into northern Europe occurred from a sub-Saharan source. BTV8 was first detected near Maastricht in The Netherlands in August 2006 and spread along a broadly east-west axis to involve most of the country, virtually the whole of Belgium, much of North West Germany, Luxembourg and northern areas of France(2). After a pause during the 2006-2007 winter, the virus continued to spread with increased vigour across northern Europe, involving many additional areas, including England, Denmark, Sweden, Norway, the Czech Republic, Poland and Hungary. The occurrence of BTV8 in Norway in late
2008 at approximately 60°N is the most northerly record for BTV anywhere in the world (see Figure 8.1 for a map of the bluetongue serotypes in Europe in early 2009).
HOST FACTORS
BTV is probably able to infect all species of wild and domestic ruminant. Severe disease usually occurs only in high-yielding domestic breeds of sheep and some species of New World wild ruminant (e.g. white-tailed deer ( Odo- coileus virginianus), black-tailed deer (O. hemionus), pronghorn antelope (Antilocapra americana), bighorn sheep (Ovis canadensis)). This means that the vast majority of BTV infections are subclinical. This covert presence of the virus, alternating with outbreaks of severe disease, has had a considerable and adverse effect upon international trade in bovines and ovines. The effects of BTV on wild European ruminants have not been investigated in detail,
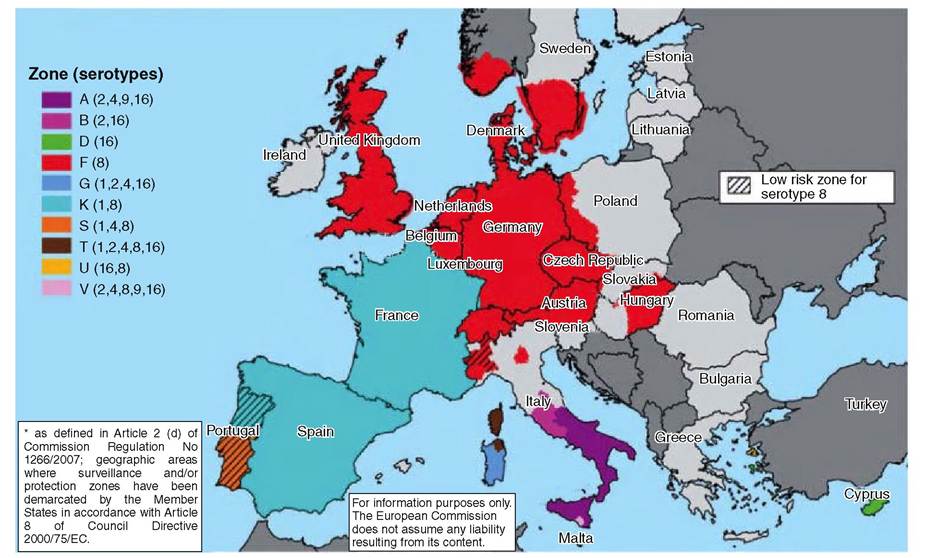
FIGURE 8.1 The European bluetongue virus restriction zones and the geographical locations of the different serotypes in early 2009.
although there are recent reports of BTV antibodies and/ or viral RNA in red deer (Cervus elaphus), fallow deer (Dama dama), Ibex (Capra pyrenaica hispanica) and mouflon in Spain(5), red deer, mouflon and roe deer in Germany1-6) and red deer in Belgium1-7).
Until recently BTV seemed to be a cause of disease in wild ruminants only in New World species. However, during the recent incursion of BTV8 into Europe clinical disease was seen in 62 captive animals originating from Europe, Asia and the Americas, and belonging to three ruminant families; Bovidae, Cervidae and Camelidae. The Bovidae seemed to be most susceptible, with four species (American bison (Bison bison), European bison (B. bonasus), Siberian ibex (Capra sibirica) and musk ox (Ovibos moschatus)) showing high morbidity and a mortality of greater than 10%. None of over 200 African ruminants of 20 species were clinically affected(8).
Antibodies to BTV-8 were identified in 40.4% of free- ranging red deer in Belgium but with no evidence of disease or increased mortality(7).
In disease-susceptible ruminant species, there is no particular age- related difference in severity of clinical signs, but these do vary between species, breeds and individuals, and depending upon the serotype/strain of the infecting virus, and certain ill-defined interactions with the environment — for example, stress and strong sunlight exacerbate disease signs.
There is no sex-related difference in the extent and severity of clinical signs in domestic ruminant species, but transplacental transmission may occur, particularly with cell culture - adapted or live vaccine viruses. In such instances resorption, abortion, birth of weak or deformed offspring, or the birth of viraemic offspring may result. The northern European strain of BTV8 is unusual as transplacental transmission in cattle seems to be astonishingly common and rates in excess of 30% have been reported1-9). The reasons for this are uncertain, but this strain of BTV8 originated from sub-Saharan Africa, a region where live BTV vaccines have been used for decades. It is possible that this BTV8 strain has acquired one or more genome segments from a vaccine virus, by reassortment, enhancing its ability to cross the placental barrier. Such reassortants have been demonstrated in the field in Europe(10).
ENVIRONMENTAL FACTORS
Vectors and Transmission
BTV is an arbovirus and so is transmitted between its hosts almost entirely by the bites of vector species of arthropod, which are certain species of Culicoides biting midge. The distribution of BTV is therefore limited to those regions where vector species occur and its transmission to those times of the year when climatic conditions are favourable for adult vector activity and for virus replication in, and transmission by, them.
In most epidemic zones BT is usually seen in late summer and autumn, as this is when high numbers of adult vectors occur and when the temperature is suitable for transmission.
In such situations the annual bouts of disease may reflect new virus introductions or low-level persistence. Reintroduction is possible if endemic viral foci are close by, as infected Culicoides may be transported on the wind over distances in excess of 100km(11). As BTV has not been proven to be transmitted vertically through its vectors and is rarely transmitted directly from vertebrate to vertebrate, except in the case of transplacental transmission, long- term persistence usually occurs only in areas where adult vectors are present virtually throughout the year. If vector- free periods (VFP) do occur, persistence may still be possible, if they are shorter than the maximum period of viraemia in the local ruminant population (up to 54 days in sheep and 60 or more days in cattle), otherwise the last infected host will have died or recovered before new vectors emerge. The part played by wild ruminants in such overwintering is uncertain, as the maximum duration of viraemia has not been determined for any wild species.BTV Vector Species of Culicoides in Europe and Climatic Effects on Vectors
Culicoides imicola is an Afro-Asiatic species and has long been known as a BTV vector in those parts of the Mediterranean Basin affected before 1998. The 1998—2005 BTV incursions into many parts of the Mediterranean Basin previously unaffected, including areas beyond the known range of C. imicola, was an unexpected turn of events. This suggested either that the range of C. imicola had expanded or that other species of Culicoides were, for the first time, transmitting the virus. In the event, the answer seems to include both these possibilities. Surveys in the BTV- affected areas from 2000 recorded C. imicola in many regions where it had been looked for in the 1970s and 80s but not found, i.e. mainland Greece, mainland Italy, Sicily, Sardinia, Corsica, the Balearics, eastern Spain and parts of southern mainland France(2). The range of this species, therefore, seems to have expanded northwards, in recent times, to include much of the northern coast of the Mediterranean Sea and most of peninsular Italy.
It has been suggested that this expansion is being driven by ongoing changes in European climate, which made the 1990s (i.e. the period when BTV incursions commenced) the warmest decade on record, and that extensions in the range of C. imicola have been into those areas that have warmed the most(12).However, BTV has spread beyond the new range of this vector(2), and even within the overall distribution of C. imicola, transmission has occurred in locations where it is either rare or absent1-12’13). This means that in these areas indigenous European vector species of Culicoides must be transmitting the virus. Recent studies have shown that two widespread and abundant Palaearctic Culicoides species complexes (C. obsoletus plus C. dewulfi and C. chiopterus, and C. pulicaris) are the likely vectors1-2). Earlier studies suggested that these species had very low oral-susceptibility rates for BTV(14). However, the recent climate warming in Europe is likely to have increased their importance as vectors, both by increasing their population sizes and by enhancing their susceptibility to BTV infection through temperature-controlled virus developmental effects1-15).
The role of these indigenous vectors in the transmission of BTV was further confirmed in 2006, when an incursion of BTV8 occurred directly into northern Europe. Despite the absence of C. imicola, the virus was transmitted widely during 2006-2008(2).
EPIDEMIOLOGICAL ROLE OF THE VERTEBRATE HOST SPECIES
Historically, it is likely that the primary BTV cycle involved wild African ruminants and Culicoides midges. The virus now seems to be maintained in a midge-cattle cycle and, once a certain level of infection is attained, spills over to initiate a secondary cycle in sheep. This generally occurs in late summer or autumn when vector populations are maximal. Cattle are the preferred hosts of vector midges and consequently are more frequently infected than are sheep; however, they rarely develop clinical disease but do exhibit an extended viraemia and are therefore an ideal virus reservoir. In essence, sheep are the tip of the iceberg and are where disease is usually seen; cattle are the covert reservoir where the vast majority of the virus resides.
Little information exists on the role, in BT epidemiology, of free- living wild ruminants in Europe. However, such data that do exist suggest that during a BTV outbreak the larger Cervidae are regularly infected and, in the absence of clinical disease, may act as covert reservoirs of infection for domestic livestock. In Spain, BTV mean seroprevalences in red deer of up 21.9% (309/1409), in fallow deer (Dama dama) of 35.4% (34/96), in roe deer (Capreolus capreolus) of 5.1% (2/39), in mouflon ( Ovis aries) of 13.2% (9/68) and Barbary sheep (Ammotragus lervia) of 25% (1/4) were recorded (16). Similarly, in Belgium seroprevalence rates of up to 40.4% were recorded in red deer(7). Also in Spain, recent work has shown that BTV RNA is able to be detected by reverse transcription polymerase chain reaction (RT-PCR) in the blood of red deer experimentally infected with BTV1 and BTV8, for periods of up to 112 days post-infection (dpi)(17). However, as has been demonstrated by many researchers, the mere presence of viral RNA has no relevance to transmission. It is therefore important that this Spanish work is followed up by elucidation of the duration and titre profile of ‘ live virus’ viraemia in common European deer species so that the potential for these animals to act as BTV reservoirs can be accurately assessed.
I nterestingly, limited data suggest that BTV may also be able to infect some carnivore species. Serological evidence from Africa suggests that large carnivores may be infected, although smaller species are not. The data indicate that the large carnivores are infected via an oral route, presumably through feeding on BTV-infected rumi- nants(18). Also, in North America the use of a canine vaccine contaminated with BTV caused abortion and death in pregnant bitches. In neither of these cases is it known whether a viraemia develops subsequent to infection or whether vector Culicoides habitually feed upon large carnivores. Consequently, currently there is no evidence to suggest that dogs or other carnivores are important in the natural transmission of BTV.
More on the topic EPIDEMIOLOGY:
- Molecular Epidemiology of Bovine Tuberculosis in Uganda
- The Epidemiology of BTB in Malawi
- Prevalence and Epidemiology of BTB in Rwanda
- Conclusion and Recommendations
- Policy Provisions for Bovine and Zoonotic Tuberculosis in Uganda
- AVIAN CHOLERA
- Cattle Movement Networks in Uganda
- Resuscitation of MAP and Growth Stimulation
- Bovine Tuberculosis in Uganda
- Voluntary counselling and HIV testing