EPIDEMIOLOGY
geographical DISTRIBUTION and HOSTS
The known distribution of LB in Europe is closely tied to the presence of ticks of the Ixodes ricinus complex, i.e. I. ricinus in most of Europe and both I.
ricinus and I. per- sulcatus in some locations in Finland, the Baltic states and parts of Russia. Ixodes ricinus typically has a restricted distribution range in sites with a Mediterranean component, such as Greece, Italy, Spain, Portugal, France and northern Africa, but is more common in Continental Europe, the UK and southern/central Sweden and Finland. North of that, and in Norway, the tick is typically restricted to the coastline, but may be found in some isolated locations as far north as 68-70°N(19). Within its distribution range, I. ricinus is most prevalent in areas where it can find protection from desiccation. This tick needs a relative humidity above 80% to survive and is found in areas with a wet and cool climate or where the microhabitat provide sufficient humidity (i.e. low vegetation, leaf litter, thick moss, grass or heather). This may typically be deciduous and mixed forest, but also other habitats where sufficient relative humidity allows tick survival. The tick population density generally decreases with altitude(20).The prevalence of different genospecies of B. burgdorferi sl (and probably also the different genotypes within each species) seem to vary greatly between regions. Borrelia afzelii and B. garinii are the most common species in most of Europe, followed by B. valaisiana and B. burgdorfers ss. Borrelia lusitaniae is rare or absent in most regions, but prevalent in south-western Europe(21) and is the sole species of LBS in southern Portugal and North Africa. Borrelia spielmaniii and B. bissettii also seem to be found only in low prevalences and have a limited geographical distribution, and the most recently proposed species, ‘B.
bavarien- sis’ seems to have a highly focal distribution1-22). The differences in prevalence of the different genospecies could probably be explained by the prevalence of different reservoir hosts.In addition to the borrelia cycle associated with I. ricinus, I. uriae transmit B. garinii among seabirds(23), whereas I. hexagonus transmit several LBS among hedgehogs, mustelids, foxes and other medium-sized animals1-24). Both these ticks are nidicolous, meaning that they live in the nest or burrow of their main host, and only occasionally bite humans and domestic animals. These semiindependent cycles of transmission may hence constitute ‘silent foci’ of B. burgdorferi in areas where I. ricinus is absent, meaning that LBS are present in the environment, even when human cases are rarely diagnosed. This implies that LBS infection in wildlife may be found further north, at higher latitude and perhaps even in dryer areas than commonly thought.
HOST FACTORS
Ixodes ricinus and I. persulcatus feed on virtually any mammal, bird or reptile, but only some of these species acquire systemic infection with LBS, which is determined to a large extent by the host’s innate immune response. In ticks feeding on host species resistant to a strain of LBS, the complement pathways will activate C3b, which binds to the surface of the bacteria, inducing formation of the terminal complement complex and hence lysis of the spi- rochaetes already in the midgut of the tick(25). However, some strains express complement regulatory-acquiring surface proteins (CRASP) on their surface that specifically bind complement-control proteins of a given animal species. This mechanism is proposed to prevent lysis. Typically, B. garinii and B. valaisiana will be lysed in the midgut of a tick feeding on rodents, whereas B. afzelif will manage to survive exposure to rodent complement. In ticks feeding on a thrush, pheasant or seabird, however,
B.
garinii and B. valaisiana will survive and B. afzelif will get killed, whereas exposure to cervid blood seems to induce lysis of all genospecies.When an infection is established, the acquired immune response may be decisive for the outcome. Young animals and immunocompromised individuals have a greater risk of widespread bacterial dissemination. Experimental infection of neonates, for example, causes arthritis and carditis not seen in adults of the same species(26).
Tick infestation loads (and thereby borrelia infection risk) may be linked to characteristics of both the host species and the individual of a species. Some species have the ability to mount an efficient immune response against Ixodes ticks, thereby presumably preventing borrelial infection. This may be the case for voles, but not for mice(27). Similar mechanisms may be at play in birds, where ticks feeding on a bird species such as the dunnock (Prunella modularis) have been found to be starving and having very low prevalence of infection compared with ticks on other ground-feeding birds(28). Other species may be able to mechanically remove a significant proportion of the ticks from their skin(29).
Among rodents, males often have higher tick loads than females. This could be explained by differences in the level of testosterone, affecting both innate and acquired resistance to tick feeding, but also by sexual differences in spacing behaviour and size(27). Space use and tick load may also be related to the individual behaviour of the animal(30).
ENVIRONMENTAL FACTORS
Borrelia burgdorferi sl is maintained in transmission cycles between Ixodid ticks and a wide variety of vertebrate hosts. Isolation or DNA detection of B. burgdorferi sl has also been described from several other arthropods, including mosquitoes, tabanid flies and fleas, but so far the Ixodidae seem to be the principal group of hosts. In Europe, the most important vector, at least for human LB, is I.
ricinuf. However, natural infections by B. burgdorferi sl have been recorded in 25 other ixodid tick species, and vectorial competence has been confirmed experimentally for 12 of these species(16).The environment for B. burgdorferi sl consists of ticks and vertebrate hosts. The biotic and abiotic factors that are decisive for the tick population, the vertebrate host population and the interaction between them determine the fate of the population of LBS. To understand the ecology of these bacteria, one has to understand the ecology of ticks (see (17) for a thorough review)(27). The I. ricinus tick has a complex life cycle, including four developmental stages: the egg and the three instars — the larva, the nymph and the adult (Figure 27.1). All three instars rely on blood as their only food source, and blood-feeding initiates development to the next life stage or, for the adults, reproductive activity. The life cycle of a European I. ricinuf may take from 3 to 6 years to complete. Most of this time, the tick survives sheltered in the vegetation litter of the forest floor. In its periods of activity, it searches for a host by questing, i.e. sitting and waiting for an animal to pass by, to which it can cling and feed on. The feeding phases,
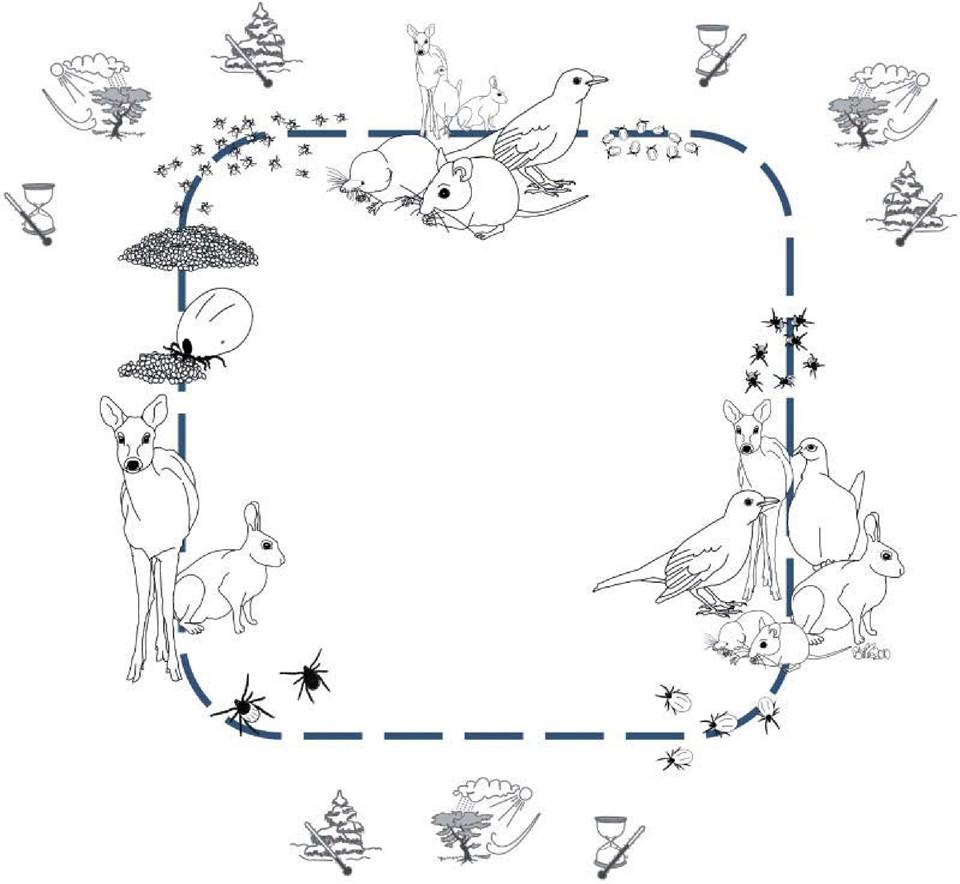
FIGURE 27.1 The life cycle of I. ricinus tick and factors that determine tick population density. The infection pressure of Borrelia burgdorferi s.l. on a given species, in a given locality, depends on a complex range of factors: firstly, on the abundance of ticks and the degree to which they feed on hosts competent to transmit the spirochaete to that species — the tick population density will depend on the suitability of the microclimate (illustrated by the tree, cloud, sun and wind above), i.e. to which degree the environment is humid enough and sufficiently protected against desiccation; the length of the growth season (illustrated by the hourglass and thermometer), i.e.
how long the temperature is high enough to allow both development from one stage to another and host-seeking activity; protection against cold-stress during winter (illustrated by the tnow-covered spruce and thermometer); and a high density of suitable hosts (illustrated by the various host animalt above).consequently, only occupy a minor proportion of the life of a tick.
The survival of the ticks, and the tick population density, may roughly be said to depend on five factors (Figure 27.1):
1. The presence of a suitable microclimate: ticks are very susceptible to desiccation caused by low relative humidity, sun and wind. Frequent rain and mist, or vegetation that shields from sun and wind, creates a humid microclimate that increases survival.
2. Long enough growth season to permit successful host seeking and development: little or no development takes place between 7 and 10°C, whereas the rate of development seems to increase with temperatures above that. Host-seeking activity by the tick larvae does not occur below a threshold temperature of 7—10°C, and for nymphs and adults does not occur below 5—7°C. Consequently, high temperatures over long periods are beneficial for the ticks.
3. Relatively mild winter or protection against very cold temperatures: on the one hand, temperatures below —10°C seem to severely diminish the tick population density, whereas deep snow cover, on the other hand, may provide effective insulation and ensure survival.
4. Presence of sufficient numbers of hosts to facilitate succession from the larval and nymphal stages: I. ricinus is an opportunistic parasite that clings to whatever animal passes by, but as larvae quest (sit and wait for a host) on lower vegetation than nymphs, a larger proportion of this instar feed on small mammals and ground-feeding birds. The population density of these animals will be vital, not only to allow development into the next instar, but also to minimize the questing time and thereby the risk of desiccation and energy depletion.
5. Presence of sufficient numbers of a large animal hosts, facilitating production of high numbers of eggs by the adult female tick.
FACTORS THAT AFFECT PREVALENCE
AMONG TICKS
Although tick population density is a major determinant of LBS infection pressure, one should not expect a strict linear relationship between tick abundance and incidence of borrelia infection in hosts. The transmission cycles for B. burgdorferi sl, having a long-living systemic infection, are so robust that more or less wherever populations of competent ticks and hosts exist, the pathogen transmission cycle will be maintained. However, as transovarial transmission is rare, and different hosts have different ability to transmit the bacteria, the prevalence of LBS among the ticks may vary with temporal and spatial variations in host composition.
Furthermore, recent studies on I. scapularis and its hosts in North America suggest that abiotic factors such as daylength and a climate- related degree of temporal synchrony between larval and nymphal questing behavior may be an important determinant both for the overall prevalence of B. burgdorferi sl in ticks and the geographical variation in prevalence of different strains(31). This model suggests that more or less seasonal synchrony of the immature instars will affect the efficiency with which ticks acquire infection from infected hosts, the duration of host infectivity relative to the timing of tick activity and the capacity for co-feeding transmission. Consequently, abiotic factors such as daylength and climate may be important determinants for the prevalence of LBS in ticks and hosts, the prevalence of different strains, and the pathogenicity of these strains.
EPIDEMIOLOGICAL ROLE OF
THE AFFECTED SPECIES
To be a reservoir host of LBS, a vertebrate must: i) harbour vector ticks; ii) be susceptible for infection; iii) allow multiplication, development and persistence of the spirocha- etes; and iv) allow transmission back to subsequently feeding ticks without diminishing the ticks’ chances of successful development and feeding in their next life stage(16). Different host species show different degrees of susceptibility for infection with different genospecies (or even strains) of LBS and consequently also varying degree of competence to support the transmission of LBS. The overall prevalence of infected ticks and the proportional prevalence of the different strains of B. burgdorferi sl in a given location may be a reflection of the proportional abundance of tick host species in that area, the relative proportion of the tick population that feeds on each host species and the degree of transmission competence each of these has for the actual strains.
Birds have a critical role as reservoirs ofLBS. In terrestrial ecosystems especially, the species that forage on the ground in forest habitats are frequently infested with I. ricinus larvae and nymphs. Some of these species are also competent reservoirs for B. garinii and B. valaisiana, though the full avian reservoir range remains to be defined. Among the relatively few species investigated, thrushes (Turdus spp.) and the ring-necked pheasant (Phasianus colchicus) seem to play key roles in the maintenance of the terrestrial cycle of these two genospecies(28’32). It has also been suggested that passerine birds function as reservoirs of B. lusitaniae(33). In addition to the role of some species as competent reservoirs for Borrelia, migratory birds are important for dispersal of infected ticks to new areas, where local reservoir hosts can contribute to the establishment of new foci for the transported borrelial strain. The function of the bird may theoretically be that of a mechanical vector, but it has been shown that migratory stress can reactivate latent borrelial infection(34). Migration may hence be associated with a temporally increased reservoir competence, facilitating long-distance dispersal of borrelial spirochaetes.
In marine environments, a wide variety of colonial seabirds seem to constitute competent reservoirs for B. garinii. Here, the main vector is the tick I. uriae. Closely related B. garinii sequences in ticks from localities in both polar regions point at a trans-hemispheric dispersal of this geno- species by seabirds, most likely by reactivation of latent infections(32). Ixodes uriae seem to form distinct races associated with one, or a few, closely related seabird host species (e.g. razorbills, guillemots, puffins, kittiwakes, fulmars) who tend to use different substrates within the breeding colonies, leading to the maintenance of several semi-independent disease cycles(35). Correspondingly, B. garinii strains isolated from I. uriae show great diver- sity(23). The terrestrial and marine transmission cycles of B. garinii are not totally separated, as some exchange of strains between them seem to occur. The marine cycles may hence serve as donors of new strains into the terrestrial ecosystems. Interestingly, recent studies have also detected DNA sequences grouping with B. burgdorferi ss and B. Iusitaniae in arctic birds, suggesting that transmission cycles of genospecies other than B. garinii also may be maintained by I. uriae and marine birds(35).
Rodents such as mice, voles and rats are thought to constitute important reservoir hosts for genospecies like B. afzelii, B. burgdorferi ss and ‘B. bavariensis'(36). The mice in the Apodemus genus appear to remain persistently infectious for ticks once infected. Although infection is rarely detected by isolation from the animals, they feed a substantial proportion of the larvae, supplying the ecosystem with many infected nymphs. Conversely, in the bank vole (Clethrionomysglareolus), although infection results in high levels of spirochaetes in blood, the vole may not function as an important reservoir, as this species seems to mount an immune response against the ticks that diminishes their probability of successful feeding and moulting and thereby reduces the trans-stadial transmission of Borrelia.
Edible dormice (Glis glis), hazel dormice (Muscardinus avellanarius( and garden dormice (Eliomys quercinus) appear to constitute the key reservoir hosts for B. spielmaniiiτ,, explaining the limited distribution of this genospecies.
Squirrels (Sciurus spp.) are also important reservoir hosts, as they are often heavily infested with subadult ticks, and these feeding ticks have a high prevalence of borrelial infection, mainly B. burgdorferi ss and B. afZelii∙v6l.
There is a dearth of knowledge on the role of different species of shrews (Soricidae) in perpetuation of LBS in Europe. Based on studies in a location in eastern USA, Brisson and others (2008)(38) suggested that shrews are key reservoirs of B. burgdorferi ss. Evidence for an important role of shrews, especially the common shrew (Sorex araneus) has also been found in Sweden(39).
Hedgehogs also have some degree of reservoir capacity for several genospecies of LBS. This host may in addition function as a bridge between the two enzootic terrestrial cycles mentioned above, as hedgehogs are often infested by both I. ricinus and I. hexagonus.
Hares (Lepus timidus and L. europaeus) may play an important role in borrelial ecology, as these hosts are able to function both as maintenance hosts for all instars of ticks and have some degree of transmission competence, at least for B. burgdorferi ss. Hares may hence support enzootic cycles of borrelia in the absence of other mammalian species(40). The role of rabbits ( Oryctolagus cunicu- lus) is more controversial.
The role of cervids is also much debated. It is often stated that the increase in deer populations in Europe (and North America) is the main reason for the increase in human cases of Lyme borreliosis, as cervids are important as management hosts for the adult ticks. Despite this, studies on roe deer ( Capreolus capreolus), fallow deer (Dama dama), white-tailed deer (Odocoileus virginianus), red deer (Cervus elaphus) and moose (Alces alces) suggest that these species are incompetent reservoir hosts for B. burgdor- ferft6∖ It has been suggested that although cervids increase the tick population, they serve as zooprophylactic hosts, serving to drive down the LBS infection rate in ticks. The degree of zooprophylaxis would depend on the relative proportion of the ticks fed by deer compared with competent reservoir species. So far, models suggest that zooprophylaxis only occurs at unrealistically high densities of deer(41). On the other hand, studies in which host and borrelial DNA are isolated from ticks show that infected ticks may have fed on cervids(42). Together with studies describing dermal infection with several genospecies of B. burgdorferi sl in cervids(43), this may suggest that co-feeding infection (see below) may occur in European cervids in the same manner as it does in Japanese sika deer (Cervus nippon) and sheep(44,45).
Little is known about the role of other large mammals such as badgers (Meles meles), other mustelids, foxes ( Vulpes spp.), wild boar (Sus scrofa) and bovids in the perpetuation of LBS. The presence of borrelial DNA from many of these species, generally not considered as reservoir hosts, may indicate that they to some degree could serve as reservoirs, perhaps through co-feeding transmission1-42).
Reptiles have been regarded as refractory for LBS infection. However, several species of European wall lizards (Lacertidae) have now been identified as key reservoir hosts of B. Iusitaniae.
TRANSMISSION
Transmission of B. burgdorferi sl. between ticks occurs mainly from infected nymphal ticks of one cohort to naive larval ticks of another via reservoir hosts. There are few adult ticks compared with nymphs, and they tend to feed on incompetent hosts, rendering them only a minor role in the perpetuation of LBS. Likewise, the role and importance of vertical transmission seems to be minor. However, because of the high fecundity of female ticks, the contribution of vertical transmission to perpetuation of borrelia may be important in some systems.
Infection of vertebrate hosts relies entirely on transmission from ticks. There is no evidence for horizontal or vertical transmission from vertebrate to vertebrate.
I nfection occurs when a fraction of the spirochaetes residing in the midgut of the tick responds to the arrival of blood by invading the salivary glands through the haemocoel. The spirochaetes follow the saliva into the feeding pool, i.e. the small, persisting haemorrhages the tick sucks blood from, multiply and invade the surrounding dermis. From the dermis they travel through the bloodstream, and finally disseminate to skin and other target organs such as joints, nervous tissue, heart and arterial walls. The ability and efficiency of tissue invasion may vary between genospecies(13). It is hypothesized that spiro- chaetes persist in the extracellular matrix of these organs, remaining available for transmission to a subsequently feeding tick(46). When new ticks start to feed it is believed that the spirochaetes migrate towards the feeding pool, probably directed by chemotactic components of the tick saliva.
More on the topic EPIDEMIOLOGY:
- The Epidemiology of BTB in Malawi
- Molecular Epidemiology of Bovine Tuberculosis in Uganda
- Prevalence and Epidemiology of BTB in Rwanda
- Policy Provisions for Bovine and Zoonotic Tuberculosis in Uganda
- AVIAN CHOLERA
- Cattle Movement Networks in Uganda
- Bovine Tuberculosis in Uganda
- Voluntary counselling and HIV testing
- Bovine Tuberculosis (BTB) in Cattle in Zambia
- Limitations of conventional historiography