HANTAVIRUSES INFECTIONS
PAUL HEYMAN
Queen Astrid Military Hospital, Epidemiology & Biostatistics Dept, Research Laboratoryfor Vector-Borne Diseases, Brussels, Belgium
Haemorrhagic fever with renal syndrome (HFRS) first drew the attention of the medical world in 1951, when more than 3,000 United Nations troops in Korea became ill with a ‘new’ disease, which had a 10% mortality.
However, this ‘new’ disease, now known to be due to hantavirus infection, had been discovered and rediscovered several times before in many regions.Earle, and earlier Langdon-Brown, suggested that a condition similar to HFRS was present during the American civil war (1862—1863) called ‘trench nephritis’. A similar disease affected German and Allied troops during World War I (1915—1916). A detailed description of what was then called ‘war nephritis’ was published in 1918. In Korea, Manchuria and the far east of Russia the condition has probably been endemic for centuries. A report of a milder form of the disease in Finland (Puumala fever) was published in 1934. During World War II Japanese troops in Manchuria also fell victim to a similar disease, then called songo fever. In the 1950s, HFRS emerged in former Yugoslavia, Bulgaria, Hungary and Czechoslovakia.
Before 1983 hantavirus disease was referred to with different, usually local, names in Europe and Asia: haemorrhagic nephrosonephritis, epidemic haemorrhagic fever, Korean haemorrhagic fever, nephropathia epidemica, songo fever, Puumala fever, Tula fever, etc. In 1982, the Working Group on Haemorrhagic Fever with Renal Syndrome recommended using the name ‘ haemorrhagic fever with renal syndrome’ (HFRS) to describe the different conditions(1).
Infectious Diseases of Wild Mammals and Birds in Europe, First Edition. Edited by Dolores Gavier-Widen, J. Paul Duff, and Anna Meredith. © 2012 Blackwell Publishing Ltd.
Published 2012 by Blackwell Publishing Ltd.AETIOLOGY
Hantaviruses are enveloped viruses with a tri-segmented (S, M and L segment) negative-stranded RNA genome. The small (S) segment encodes the nucleocapsid, the medium (M) segment encodes two envelope glycoproteins (Gn and Gc) and the large (L) segment the RNA- dependent RNA polymerase.
Several authors consider hantaviruses to be one of the best examples of a long- term association between RNA viruses and their hosts. The strong host specificity seems to suggest that hantaviruses co-speciated with the rodents and insectivores they infect, as these mammals last shared a common ancestor approximately 100 million years ago. However, there is scant evidence for a history of codivergence between hantaviruses and their hosts. Moreover, the divergence times for the hantavirus genus are several orders of magnitude more recent compared with the speciation timescale of their hosts. The phylogenetical similarities between the hantaviruses and their hosts are thus probably the result of a more recent history of adaptation and host switching. The genus Hantavirus consists of at least four serogroups or clades, with at least 40 different serotypes. As only about 10% of the more than 2,000 existing rodent species have been examined for their hantavirus carrier status, it is highly probable that the genus Hantavirus consists of 300 to 400 serotypes. Of the 40 serotypes known at present, at least 13 viruses can cause one of two major clinical syndromes, sometimes with overlapping features, in humans: HFRS in Europe and Asia, and hantavirus cardiopulmonary syndrome (HCPS) in the Americas. HFRS is caused by Hantaan, Seoul, Dobrava, Saaremaa and Puumala viruses (HTNV, SEOV, DOBV, SAAV and PUUV, respectively). SAAV and PUUV cause a mild form of HFRS referred to as nephropathia epidemica (NE). HCPS is caused by Sin Nombre, Black Creek Canal, Bayou, New York 1, Juquitiba and Andes viruses (SNV, BCCV, BAYV, NYV, JUQV and ANDV, respectively).
EPIDEMIOLOGY
GEOGRAPHICAL DISTRIBUTION
PUUV, Tula (TULV), DOBV, SAAV and SEOV hantaviruses have been reported as circulating in Europe. NE is typical in PUUV and SAAV infections. HFRS is typical for HTNV, DOBV and SEOV infections. PUUV and DOBV have caused the vast majority of human cases in Europe(2) (Figure 18.1). SAAV has also recently been found to be responsible for human cases in Eastern Europe. TULV has been linked to human infections in the Czech Republic, Switzerland and Germany. No confirmed human cases of SEOV infection have been reported in Europe so far, although an unpublished case, confirmed by focus reduction neutralization tests (FRNT), occurred in France. In Western and Northern Europe only PUUV infections have been reported. From west to east across Europe, the DOBV infection prevalence in humans varies from 3.6% in Southern Germany, to more than 50% in Slovenia and 100% in Greece.
During the past two decades, the number of diagnosed human cases has been increasing constantly in almost all European countries. Since 2002, record case counts have been noted in Finland (2,603 cases in 2002, 2,526 cases in 2005), Sweden (459 cases in 2004), Germany (more than 1,400 cases reported in September 2007) and Belgium (372 cases in 2005). Asymptomatic or mild infections that go unnoticed probably result in an underestimation of the number of hantavirus infections. The ratio ranges of sub- clinical to clinical infections for PUUV infection is from 5:1 to 10:1 in Europe; it is thus reasonable to assume that the actual number of infected individuals is 5 to 10 times higher than the reported cases. An 8-year study in Finland demonstrated that the estimated ratio of diagnosed HFRS cases was only 13% (4 to 30% for different areas), leaving at least 70% of the PUUV infections undiagnosed because they were subclinical or showed only minimal or atypical symptoms. PUUV, DOBV and HTNV infections are predominant in rural areas, whereas SEOV infections occur in urban settings.
Investigations have linked handling wood, farm work, sleeping on the ground, military exercises and lower socio-economic status to increased risk for acquiring hantavirus infection. HFRS is more common in males than in females (the male : female ratio can reach 2:1 to 3:1), and most of the cases occur in the 20—50 age group. In most European countries the seroprevalence rate in the general population ranges from 1 to 5%, with peaks in certain regions (Finland, Bosnia Herzegovina) up to 20% (Figure 18.1).ENVIRONMENTAL FACTORS
The distribution of hantavirus infections in time and space reflects the distribution range and population densities of
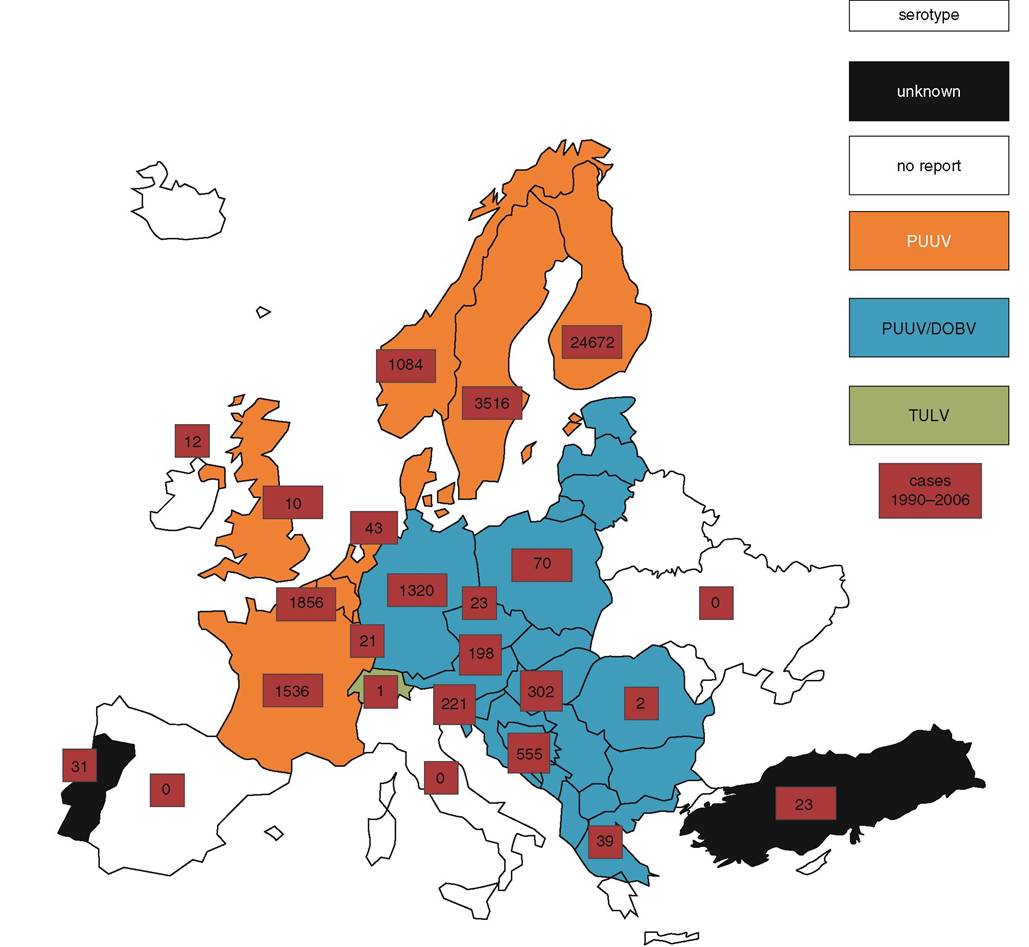
FIGURE 18.1 Human hantavirus cases in Europe (1990—2006).
their rodent reservoir host vector. High reservoir host abundance induces higher transmission rates to humans. In Northern Europe the bank vole (Myodes glareolus) population frequently has a 3—4 year cycle of abundance, which coincides with the PUUV infection patterns in the respective regions. These cycles are based on a predator-prey mechanism. In Western Europe, high bank vole population density is related to high tree seed production (mast years), which is triggered by particular climatic conditions. In Belgium, it was demonstrated that NE epidemics were preceded by abundant tree seed production, which in turn was directly linked to climate(3). The increased hantavirus incidence in humans was preceded by high summer temperatures 2 years earlier and high autumn temperatures 1 year earlier(4).
EPIDEMIOLOGICAL ROLE OF THE AFFECTED SPECIES
Rodents and insectivores act as hantavirus vectors. The viruses are found in the Cricetidae family (subfamilies Arvicolinae, Neotominae and Sigmodontinae) and in the Muridae family (subfamily Murinae). Each hantavirus type is carried by a specific rodent host species, and phylogenetic analysis has revealed that the relationships between hantaviruses parallel the phylogeny of their rodent hosts, although host switches have occurred.
Recently, new hantaviruses have been identified in insectivores. It is noteworthy that Thottapalayam virus (TPMV), which is carried by an insectivore, Suncus murinus (the Asian house shrew, or brown musk shrew), first described in 1971(5) precedes HTNV, which is carried by Apodemus agrarius (the striped field mouse) and was first described in 1976(6), although the latter was designated as the prototype virus. In nature, hantaviruses are maintained by horizontal transmission (rodent to rodent, via fighting, biting or grooming) and via aerosolized contaminated excreta.In Europe, HFRS is caused by four hantaviruses: PUUV, carried by Myodes glareolus (bank vole); DOBV, carried by Apodemus flavicollis (yellow-necked mouse); TULV, carried by Microtus arvalis (common vole); and SAAV, carried by A. agrarius (the striped field mouse). In Asia, HFRS is caused by HNTV and the Amur virus (AMRV), which are transmitted by A. agrarius (striped field mouse) and A. peninsulae (Korean field mouse), respectively, and by SEOV, which is transmitted by Rattus rattus and R nor- vegicus (the black rat and the brown rat). Although R norvegicus in Europe was found to carry SEOV, the virus does not seem to be a public health problem at present.
A clear negative effect of high species diversity on both abundance of reservoir hosts and their infection rate with hantaviruses in Panama, South America, was demon- strated(7). However, a different pattern seems to exist for Sin Nombre hantavirus (SNV) in deer mice (Peromyscus maniculatus) in North America, where a negative correlation between habitat disturbance and abundance and seroprevalence was noted. In Belgium, a correlation between high host diversity and low rates of pathogen transmission, or disease risk, for PUUV was noted(3). These findings strongly suggest that high biodiversity contributes to the protection of human health.
All non-rodent mammals, with seropositive populations, are considered dead-end hosts, i.e.
not capable of transmitting the infection to conspecifics. Little is known, however, about the presence ofhantaviruses in non- rodent mammals. Experimental infection in various non-human primates has led to an antibody response1-8). Antibodies to hantaviruses were detected in domestic cats (Felis catus)9 and dogs (Canisfamiliaris)1-10), and in red foxes ( Vulpes vulpes)^. In Belgian dogs and cats, a seroprevalence of 4.9% and 16.9%, respectively, was found. In Sweden, antibodies to PUUV were detected in moose (Alces alces) from endemic areas(12).Coyotes (Canis latrans), island foxes ( Urocyon littoralis) and 25 individuals from seven different mammalian species were all found negative for hantaviruses in California1-13). HTNV was detected in a bird, E mberiza elegans, in Russia(14). Antibodies to HTNV were also detected in bats (Jung, 1995)(15) in South Korea.
The viruses are transmitted to humans by aerosol of infected urine, saliva and faecal material from rodents. In the reservoir species, horizontal transmission occurs, whereas humans and other mammals are dead-end hosts.
PATHOGENESIS, PATHOLOGY AND IMMUNITY
The infectious dose for all hantavirus serotypes is unknown, but it is considered to be low. Presumably, high viral loads cause a more severe infection. Humans become infected when they come into contact with rodents or inhale aerosolized particles. For all but one serotype (the South American ANDV) human-to-human transmission has not been described. Rodent bites may also be a route of infection, but this is rare compared with the inhalation route. Infection via the digestive tract is highly unlikely and has not been reported.
Infected rodents have life- long shedding of virus in saliva, urine and faeces, but the virus shedding is greatest approximately 3 to 8 weeks after initial infection(16). The virus is also readily detectable in the lungs and the kidneys, but no pathophysiological signs have been described.
In HFRS damage occurs to capillaries and small vessel walls, inducing vasodilatation and haemorrhages. The damage is most severe during the hypotensive and oliguric phases (see below), when widespread haemorrhages can occur. The kidney is usually involved, but pulmonary symptoms are also possible. Although limited amounts of immune complex can be found in the kidney, it is thought that viral destruction of cells plays a more important role than immuno-pathological mechanisms in inducing nephritis.
All hantaviruses induce neutralizing antibody production in the host, which provides life-long protection against reinfection with the same serotype. Cross-protection against infection with other hantaviruses has not been described.
CLINICAL SIGNS AND TREATMENT
Hantavirus infections in rodents and insectivores are persistent and apparently subclinical. However, it was reported that SEOV infection increases aggressive behaviour in male rats (R. norvegicus). PUUV infection was recently linked to impaired breeding success and winter survival in bank voles. In mammalian hosts — apart from humans — there are no reports of clinical illness, but the available information is scarce.
HTNV and DOBV cause the most severe forms of HFRS; PUUV, SAAV (nephropathia epidemica) and SEOV cause less severe illness.
HCPS in humans is characterized by fever, myalgia, gastrointestinal symptoms and abrupt onset of respiratory distress and hypotension. The case fatality rate (CFR) can be as high as 40%. Convalescence in all forms of HFRS and HCPS may take weeks to months.
Specific treatment for humans, apart from general supportive measures, is not available for hantavirus infection.
DIAGNOSIS
The diagnosis of HFRS based on clinical examination only is difficult, as hantavirus disease invariably starts with nonspecific flu-like symptoms. Diagnostic tests are required for confirmation.
Hantavirus-specific antibodies and antigen can be detected in rodent serum or tissue samples by serological and molecular biology tests, respectively. It is not possible to define the infecting serotype with standard serology tests, such as indirect fluorescent antibody (IFA) test, enzyme-linked immunosorbent assay (ELISA) or western blot, because of the cross-reactions between serotypes. For serological determination of the causal serotype, neutralization tests are required. Neutralization tests, however, are difficult and require at least a biosafety level (BSL) 3 environment.
In humans, hantavirus infection can also be diagnosed by reverse transcription polymerase chain reaction (RT- PCR) applied to blood samples. As the duration of the viraemia depends largely on the hantavirus serotype, RT-PCR must be applied as soon as possible after onset of symptoms. As a rule, higher viral loads are found in more severe hantavirus infections (HTNV, DOBV, SNV and ANDV infections), whereas the level and duration of the viraemia is considerably lower and shorter in PUUV infections. As a consequence, RT- PCR can only detect viral RNA in PUUV infections provided that sampling is done early enough (within the first 7 days after onset of symptoms). The detection of PUUV-RNA in the saliva of NE patients could open possibilities in diagnostic procedures. However, it must be noted that, contrary to what this finding suggests (namely the possibility of person-to-person transmission by close physical contact), PUUV is not known to be transmitted this way.
MANAGEMENT, CONTROL AND REGULATIONS
As hantaviruses are carried by sylvatic rodent species, management and control of populations is hardly feasible. Currently, no nationwide rodent control programmes are in effect. Pest control in agriculture and improved habitat conditions for the natural predators of rodents (mustelids, birds of prey, foxes, feral cats, etc.) may have beneficial effects.
Safe and effective vaccines are needed urgently in order to reduce the incidence of human illness, but to date these are not yet available in Europe or the USA, although M-segment DNA vaccines to HTNV and PUUV are in preparation. A vaccine against HTNV grown in suckling mice brain has been shown to induce protein immunity in mice and humans. It is currently available under the name Hantavax® and is licensed for use in the Republic of Korea, but neither EU nor FDA approval was granted.
Hantaviruses, like most enveloped RNA viruses, are susceptible to 1% sodium hypochlorite (10% sodium hypochlorite for heavily soiled material), 70% ethanol or 2% glutaraldehyde. Infected Vero E6 cells are completely inactivated after heating for 1 hour at 60°C, while about 90% of infectious particles are lost by heating at 56°C for 30 minutes; HTNV has shown sensitivity to a pH of 5. Infectivity of hantaviruses has been reported to persist in neutral solutions for several hours at 37°C and for up to 10 days at lower temperatures. Dried cell cultures remain infectious for up to 2 days. Virus suspensions stored at —60°C in 1% bovine albumin salt solution remain infectious for over 5 years. Recently, it was shown that PUUV can survive outside the host for at least 10 days at room temperature and probably much longer (weeks or months) at lower temperatures — and, for instance, under snow cover.
Hantavirus disease is a notifiable disease in the majority of European countries; it is not notifiable in Austria, Cyprus, France, Portugal, Romania and Spain. In some countries, mainly in Eastern Europe, the legally required reporting status is not yet determined®.
PUBLIC HEALTH CONCERN
Hantaviruses are globally important pathogens and cause thousands of human cases each year in Europe, thus having a significant public health impact. Depending on the serotype, the CFR may be high. In Europe the CFR for PUUV is to occur (endemic activity), but widespread rains, allowing large populations of mosquitoes to develop, may be necessary for widespread epidemics. In areas such as the Nile Delta and the Arabian Peninsula, the absence of mosquitoes species in which transovarial transmission of the virus can occur may explain why the virus has not apparently become endemic.
There is no compelling evidence identifying any one African species of mammal as the natural reservoir of RVF virus. (Indeed, there is arguably no need for a mammalian reservoir host in areas where the virus is vertically transmitted within floodwater mosquitoes and can lie dormant in egg rafts for many years.) Serological surveys of wildlife indicate that several species of ruminants, carnivores and bats can become infected with RVF. Once climatic conditions are favourable for the hatching of the RVF-infected eggs of floodwater species, the African Cape Buffalo (Syn- cerus coffer) may act as an amplifying host, in a similar way to domestic ruminants.
Adult ruminants develop extremely high levels of viraemia, which allow many species of mosquito to be biological vectors. When climatic conditions are favourable for large mosquito populations to build, epidemics can occur over an extensive area, sometimes involving several neighbouring countries.
In domestic ruminants, RVF is predominantly a disease of sheep and goats, characterized by abortions and neonatal death. It is not unusual to see in excess of 90% of newborn lambs dying of RVF during an epidemic. The disease is characterized in domestic livestock by the sudden onset of lethargy, inappetence, high fever, nasal discharge, and diarrhoea. In the acute stages of the disease, viraemia may exceed 106 plaque-forming units (PFU) per ml. Necropsy reveals a diffuse hepatic necrosis, splenomegaly and gastrointestinal haemorrhage. Viral antigen is readily detectable in the reticulo-endothelial system and in multiple organs, including the liver, kidneys, adrenal glands, gastrointestinal tract, brain, ovaries and endometrium.
The disease in humans is usually characterized only by fever, but on occasion, by encephalitis, retinitis and/or generalized haemorrhage. Although some human cases almost certainly arise from mosquito bites, RVF is seen most frequently in individuals who are exposed to viraemic blood during the slaughter of animals and the butchering of meat. Veterinarians are at greater risk of acquiring RVF than the normal population, from drawing blood and conducting necropsies.
A comprehensive bibliography on Rift Valley fever may be found in Bird et al. (2009)(17), and specifically in relation to wildlife in Evans et al. (2008)(18).
More on the topic HANTAVIRUSES INFECTIONS:
- HANTAVIRUSES INFECTIONS
- Hantavirus Infection
- Can Species Diversity Suppress Human Diseases? A Case Study
- CRIMEAN-CONGO HAEMORRHAGIC FEVER
- SUBJECT INDEX
- 5 Appendices
- BIBLIOGRAPHY FOR VIRAL INFECTIONS
- HELMINTH-VERTEBRATE SYSTEMS
- INTRODUCTION
- Viral Hemorrhagic Fevers