THE INSECTS
Insects are characterized as having jointed exoskeletons, clear segmentation that typically results in three distinct body regions (head, thorax, and abdomen), and, at least in the adults, three pairs of jointed legs.
Many types of insects are either winged as adults or have specific castes within the adult community that are winged (such as in ants and termites), and insects are the only arthropods that fly. Seven orders of insects are associated with diseases in wildlife, domestic animals, and humans. These include the Orders Blattaria (cockroaches), Phthiraptera (parasitic lice), Hemip- tera (true bugs), Siphonaptera (fleas), Diptera (flies), Lepidoptera (butterflies and moths), and Hymenoptera (bees, ants, and wasps) (Gullan and Cranston 2005, Durden and R. 2009).Most insects reproduce sexually, but parthenogenesis is known to occur in many groups. Some groups have even evolved gender differences resulting from whether the individual is sexually or asexually produced. For example, haploid bees are asexually produced males and diploid bees are sexually produced females. Most insects oviposit eggs, but a few species (including parasitic tsetses and hippoboscid flies) are viviparous; these species produce eggs that hatch into larvae inside the female and are then larviposited into the environment via a form of live birth. Insects can be separated into groups depending on the degree of developmental change (metamorphosis) between immature and adult stages. Some insects undergo little change, and these are grouped together as undergoing ametabolous development; but none of these is associated with diseases of wildlife. Others, including cockroaches, grasshoppers, lice, true bugs, and so on, undergo paurometabolous or incomplete development. Immatures of these species hatch from eggs as first instar nymphs, and undergo successive molts through a set number of nymphal instars (3 in lice, 5-13 in cockroaches) that increase in similarity to adults with each molt (Fig.
5.1). After a defined number of molts, sexually mature adults emerge to feed and reproduce; there is no pupal stage in species that undergo incomplete metamorphosis (Gullan and Cranston 2005, Durden and R. 2009).Paurometabolous insects are referred to as exopterygotes (exo: external, ptera: wing) because the wings develop from external wing buds. Some insects, including dragon flies and damsel flies, undergo hemimetabolous development, with more dramatic changes than occur in paurometabolous insects, due to stagespecific development of complex mouthparts or gills. However, these species are not typically associated with wildlife diseases, and are not discussed further. Finally, some insects, including the flies, bees, butterflies, and beetles, undergo a complete or holometabolous development (Fig. 5.2) (Gullan and Cranston 2005).
Holometabolous insects are also referred to as endopterygotes (endo: internal, ptera: wing) because the wing buds develop internally during metamorphosis of the pupa. The pupa typically is an immobile, apparently quiescent,
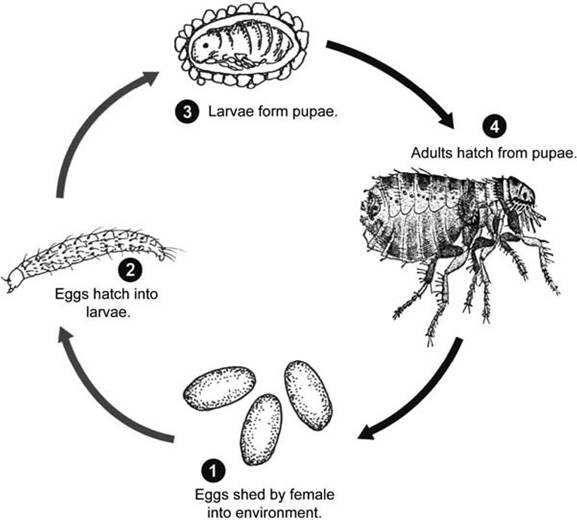
FIGURE 5.2 Life cycles of fleas, as an example of insects with holometabolous (complete) development, involve eggs, larvae (typically at least three instars), pupae, and sexual adults (Courtesy of Centers for Disease Control and Prevention's Division of Parasitic Diseases and Malaria, www.dpd.cdc.gov/dpdx).
developmental stage between the final larval stage (which looks nothing like an adult) and the adult (or imago). However, some pupas, including those of mosquitoes, are active; mosquito pupas commonly are called tumblers to differentiate them from the larval feeding stages referred to as wigglers. Nonetheless, even apparently inactive pupas may be active metabolically as they conceal the development of the imago inside. There are several types of pupas: mosquitoes and other nematoceran flies lack special protection for the pupal stage; brachyceran flies produce a puparium in which the final larval “skin” is retained as a protective case; and the fleas and moths spin silk threads to create protective cocoons.
These groups will be discussed further below under the life histories of the various groups of insects (Gullan and Cranston 2005).The degree of ecological overlap of the immature stages and the adults is another key difference in the life history strategies of those insects undergoing incomplete (pau- rometabolous) development versus complete (holometabolous) development. For examples, nymphal cockroaches, lice, and true bugs look and act like small versions of the adults, and they may compete with the adults of the same species for commonly consumed foods (Fritsche 1999, Gullan and Cranston 2005, Capinera 2010). From a disease standpoint, this allows for the additional potential for a pathogen to be picked up by one stage and transmitted by the next (e.g., transmission between successive nymphal instars or between nymphs and adults is referred to as transstadial transmission). This type of transmission is especially important in the ecology of some of the biologically transmitted pathogens and tends to stabilize enzootic transmission within a population or community of hosts.
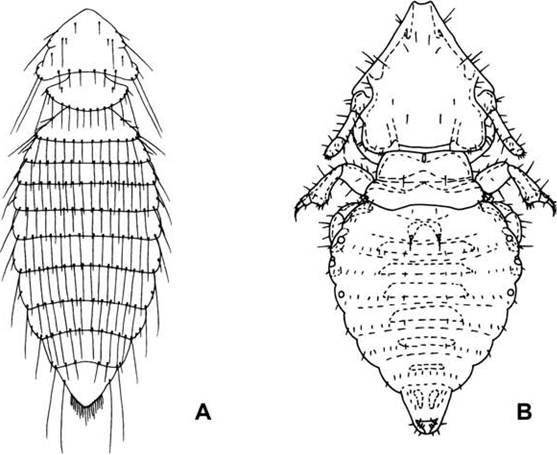
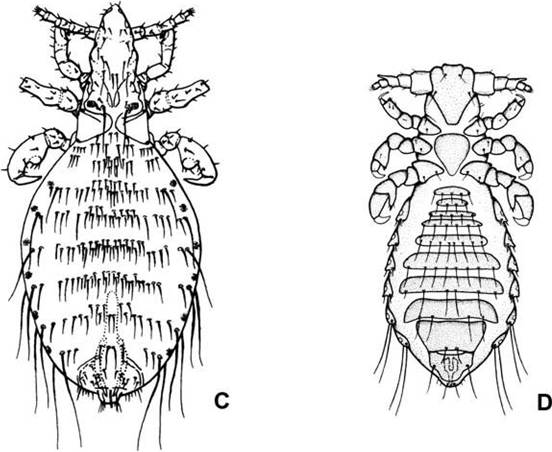
FIGURE 5.3 Representative lice found on wildlife:
(A) Menopon gallinae (Suborder Amblycera) found on game birds; (B) Felicola subrostratus (Suborder Ischnocera) found on felids, civets, and the Madagascar mongoose; (C) Linognathus africanus (Suborder Anoplura) from deer and other ruminants; and (D) Polyplax spinulosa (Suborder Anoplura) from some murid and cricetid rodents (from Capinera 2010, copyright © John Wiley and Sons, by permission).
Insects undergoing holometabolous development have larval stages that look, act, and feed very differently from the adults. Consider that worm-like butterfly caterpillars feed on leafy vegetation using chewing mouthparts while the adult (imago) butterflies tend to be delicate and sleek, and (most) feed by piercing the vascular tissues of plants.
Likewise, the maggots of calliphorid flies and the larvae of mosquitoes look nothing like the flying adults which they eventually become. Such development separates the larval forms from the adults ecologically, and tends to limit the importance of transstadial transmission of pathogens (Fritsche 1999, Gullan and Cranston 2005, Capinera 2010). In these insects, the feeding of individual adults on multiple hosts is typically necessary to achieve transmission of parasites; bird malaria (Plasmodium relictum and related spp.) is transmitted by mosquitoes that become infected by the protozoa while feeding on one host, survive long enough for parasite development (usually several weeks), and later transmit the infective stage to new, naive, hosts during subsequent blood meals.Some Insects Undergoing Paurometabolous (Incomplete) Metamorphosis
lice Lice are small, dorso-ventrally flattened, wingless (even as adults) insects in the Order Phthiraptera (Fig. 5.3). The 5,000 species of lice that feed as ectoparasites of birds or mammals were previously classified as being in one of two orders, the chewing lice (Mallophaga) and the sucking lice (Anoplura). However, recent molecular studies have demonstrated that the clade containing these groups is internal within the older order known as the Psocoptera. This leaves the order in need of revision, but we'll refer to the Order Phthiraptera. The four suborders of parasitic lice include three groups of chewing lice (the Ischnocera with approximately 1,800 species, the Amblycera with 850 species, and the Rhynchophthirina with three species) and one suborder of sucking lice (the Anoplura with approximately 550 species) (Gullan and Cranston 2005, Durden and Lloyd 2009).
The life cycle of lice is fairly simple; adults mate and females lay eggs (nits) that hatch into first instar nymphs, which undergo sequential molts into larger and more developed nymphs and then finally into the sexually mature adults. However, there are several interesting adaptations worthy of mention.
First, some species are parthenogenic, requiring no males for production of fertile eggs. Second, the genitalia of adults of non-parthenogenic species are specialized, with the male genitalia taking up nearly half of the abdomen and the female genitalia being associated with finger-like gonopods that guide, manipulate, and glue the eggs to the hair or feather shafts of their hosts. Third, the fact that the eggs are glued to the host allows generation after generation of lice to persist on a single host animal (Durden and Lloyd 2009).Lice lack the ability to move quickly and are transferred from host to host only through intimate associations (usually involving direct contact), co-habitation, and in some species (especially the Suborder Ischnocera) dispersion via phoresy. Phoresy is the process by which chewing lice, or other arthropods, attach to larger blood-sucking flies and “hitch a ride” to the next vertebrate host (Durden and Wilson 1991, Durden and Lloyd 2009). Since most species of lice are found on only one species of host, or sometimes a few related, species of hosts, pho- retic transfer is risky unless the carrier-fly is likely to feed repeatedly on the same species of host. Such close association and host specificity have resulted in phylogenetic trees of hosts that are closely mirrored by the corresponding phylogenetic trees of their lice (Hoberg et al. 1997). Co-speciation resulting from the narrow host range of some parasites (especially body lice) led to Fahrenholz's Rule (Fahrenholz 1913) which states that parasites speciate in response to differences in their hosts as a result of host specificity. While this general rule is worth remembering, parasites (and non-parasites) adapt and speciate in response to environmental pressures, some of which may be related to their hosts, to maximize individual fitness, isolation and drift, or as random genetic events (Black and Salman 2004). Thus, the cause and effect, co-evolutionary, relationships should not be extrapolated beyond what can be demonstrated quantitatively.
Host specificity of chewing lice may depend more upon their ecological niches on the hosts and their potential for phoretic transfer than to co-evolutionary history per se (Clayton and Johnson 2003). Clayton and Johnson (2003) found that avian body lice are more likely to co-speciate with their hosts, indicating specificity, than are wing-feather lice presumably because wing-feather lice are more likely to encounter larger flies and to transfer phoretically than are body lice; wing-feather lice are more likely to be relative generalists that feed on a broader range of species. In addition to host specificity, many species of lice have evolved site specificity with adaptations to characteristics of feathers or fur of specific areas of the hosts' bodies.The capacity to cause irritation or transmit disease agents varies considerably between lice that feed on blood and those that feed on feathers, skin, or hair. The sucking lice are all parasitic on placental mammals; whereas most species of chewing lice feed on birds and only about 12.4% (553 species) feed on mammals (Durden and Lloyd 2009). The Amblycera and Ischnocera can be distinguished from the Ano- plura by the relative width of the head and thorax; the heads of these chewing lice are broader than their thoraxes whereas the heads of sucking lice are narrower than their thoraxes. As suggested by the name, chewing lice have mouthparts adapted for chewing. Nonetheless, there is a gradation in adaptations from the primitive Suborder Amblycera (including Heterodoxus spiniger, a chewing louse of wild canids; Trinoton anserinum, a goose body louse; and Trinoton querquedulae, a body louse of ducks), through the Suborder Ischnocera (including feather lice of waterfowl, Anaticola spp., and the biting lice of ungulates, Bovicola spp., and canids, Trichodectes spp.) to the more specialized Rhyncophthirina (including the elephant louse, Haematomyzus elephantis). The piercing mouthparts of the Anoplura (including the genera Haematopinus, Hoplopleura, Linognathus, Solenopotes, Haematodipsus, Pedic- ulus, and Phthirus) are the most specialized of the parasitic lice.
Most populations of wildlife do not suffer population-level effects from louse infestations. However, animals that are debilitated, weakened by other diseases or parasites, or that lack the ability to groom effectively, can become overwhelmed by their populations of lice (and other ectoparasites) (Clayton et al. 2008). Heavy louse infestations are associated with considerable itching and reduced function of fur or feathers (Clayton et al. 2008, Durden and Lloyd 2009). Hosts may overgroom their fur, pick apart their feathers, or mutilate themselves when louse infestations become too great. In addition, heavy louse infestations may cause avian hosts to abandon their nests. The heaviest louse infestations seem correlated with avian host densities, and social species may be more at risk than solitary species (Clayton et al. 2008). Sucking lice cause chronic loss of blood, and heavy infestations can result in considerable blood loss, which may be especially important in young animals (Durden and Lloyd 2009). In some cases, non-native lice have been associated with deaths, and even population declines, of wildlife (refer to sections below on blood loss and dermatitis).
true bugs There are about 90,000 species of true bugs in the insect Order Hemip- tera; the name derives from the morphology of the forewings of some hemipterans; these bugs have transparent distal membranes that make them appear to have “half-wings” (hemi: half, ptera: wing). Representative hemipterans include leaf hoppers (Family Cicadelidae), backswimmers (Family Notonectidae), giant water bugs (Family Belosomatidae), kissing or assassin bugs (Family Reduviidae), bat bugs (Families Cimicidae and Polyctenidae), and the swallow and bed bugs (Family Cimicidae). True bugs have mouthparts adapted to piercing tissues, allowing them to feed on plant fluids or on other insects. Of course, some of these species (members of the Families Redu- viidae, Polyctenidae, and Cimicidae) are more specialized and have adapted lifestyles of ectoparasites that feed obligatorily on vertebrate blood. Simplistically, the life histories of this large order can be compared to other insects that undergo incomplete, or hemimetabolous, metamorphosis; eggs hatch into nymphs that undergo a series of molts (typically five) to eventually become adults that mate; females lay eggs and the cycle continues. These are generally long-lived bugs; life cycles of some species can be completed in a minimum of 24 days (bed bugs; Cimex spp.) at 30°C, but most species generally require 1-2 years at cooler temperatures (Krinsky 2009a).
Adult reduviids are winged and have large, jointed, stylette-like mouthparts. Within the Family Reduviidae, the Subfamily Triatomi- nae includes the American vectors of Chagas' disease (a zoonotic infection of Trypanosoma cruzi). These bugs live in tree holes, fallen logs, nests, roosts, burrows, caves, rock cracks, or other structures frequented by reptiles, birds, or mammals. Epidemiologically important vectors are among those species that specialize on hosts of T. cruzi, including primates and rodents. Some species are notoriously peridomestic, and it is generally these species (Rhod- nius prolixus, Triatoma infestans, Panstrongylus megistus, etc.) that transmit most of the Chagas' disease to people in South and Central America (Krinsky 2009a).
Cimicids are dorso-ventrally flattened, wingless insects. Survival within nests of seasonally breeding birds and roosts of seasonally active bats is promoted by the ability of nymphs to survive long periods (typically many months) without feeding. As with the reduviid bugs, cimicids are able to excrete fluid after feeding on a blood meal; excretion of approximately 50% of the blood meal as fluid allows ingestion of greater volumes during meals (Krinsky 2009a). Cimicids hide in substrates near nests, roosts, or burrows, as do reduviids, and leave such hiding places only to feed on host blood. As with many other ectoparasites, cimicids find their avian and mammalian hosts through recognition of higher levels of carbon dioxide and body temperatures exceeding ambient temperatures. They then congregate, sometimes in large numbers, near hosts in specific substrates that provide thermal and structural cover. Swallow bugs are considered further below in the section on blood loss (Krinsky 2009a).
Some Insects Undergoing Holometabolous (Complete) Metamorphosis
fleas The Order Siphonaptera (siphon: siphons, aptera: wingless) comprises approximately 2,500 species of highly adapted, bloodfeeding ectoparasites of mammals and birds; approximately 94% of these species are parasites of mammals. Adult fleas are readily recognized because they are laterally compressed, wingless, and have long back legs (Gage 2004, Durden and Hinkle 2009). Closer inspection allows one to appreciate the other structures that help in identification of species, including the characteristics and patterns of setae, sclerotized ctenidia (or combs of spines), sensory sensilium, and the structure of specialized genital organs. Flea structures also reflect their degree of specialization on a particular group of hosts.
Fleas have converted muscles and structures that once allowed flight in their ancestors into structures that provide an ability to jump through space, giving fleas the ability to launch themselves onto a passing host or to rapidly escape a host that is effectively grooming. The jump is not propelled by direct muscle action as might be presumed. Rather, the jump is accomplished by rapid expansion of pads comprised of a unique protein called resilin acting on the lever action associated with multiple segments of their long legs. Resilin is said to store and release energy more efficiently than rubber, and fleas can develop a jump that accelerates them to over 200 ? G through distances of greater than 30 to 50 cm (a distance of over 100 times the length of their bodies) in less than 0.02 second (Gage 2004, Durden and Hinkle 2009). For comparison, consider yourself being propelled through a kilometer of space in a fraction of a second, landing on a host the size of a skyscraper, and then digging in for a huge meal! Of course, as pointed out by Haldane (1932), such comparisons are nonsensical, but they do help to illustrate the jumping ability of fleas.
Fleas are holometabolous insects that produce larvae that look and behave differently from the adults. In fact, although most people readily recognize adult fleas, few would be expected to recognize flea larvae. Female fleas must obtain a blood meal prior to laying hundreds of small eggs, which typically fall off of the host and into the nest or burrow. Flea larvae hatch within about a week's time, and are small and worm-like, lacking legs and eyes, and their head capsules are well developed, with large mandibles adapted for chewing the organic matter found in the floor of nests or burrows. In addition to dead skin, hair, and bits of feathers, flea larvae also feed on the rich fecal pellets of adult fleas. Adult fleas excrete feces that are rich with undigested blood as they feed on the blood of their host, and the excreta provide a nourishing meal for the larvae waiting below. Each larva molts through a successive series of instars (typically three) and eventually spins a silken cocoon. The sticky silk threads used to spin the cocoon stick to bits of debris in the environment, possibly helping to camouflage the developing pupa within the cocoon. An adult emerges from the pupa after 1 to 2 weeks and is immediately ready to feed. The life cycle of some fleas can be limited to one generation per year to coincide with seasonal host availability, or, as is true for many species, they can undergo continuous generations on a single host (Gage 2004, Durden and Hinkle 2009).
Finally, fleas are important vectors of trypanosomes (including Trypanosoma nabiasi and T. lewisi, which are parasites of Old World rabbits and rodents, respectively), bacteria (including flea-borne rickettsiae and the pathogens that cause plague and tularemia), and viruses (including the myxoma virus that causes myxomatosis in rabbits). In addition, they serve as intermediate hosts of nematodes [including Acanthocheilonema (formerly Dipet- alonema) reconditum of canids] and a variety of tapeworms (including Dipylidium caninum of canids and felids and Hymenolepis spp. of rodents and primates).
flies The Order Diptera is one of the more diverse orders of insects, with over 120,000 species described (Hall and Gerhardt 2009), and is clearly the most important order of insects associated with diseases of wildlife. Whereas most flying insects have two pairs of wings, one pair each on the mesothorax (the second of three thoracic segments) and the metathorax (the third thoracic segment), the Diptera have only one pair of wings on their mesothorax. In place of the second pair of wings, flies have a pair of modified, knobbed wing buds called halteres that are used for balancing during flight. The order includes two suborders: the Nematocera includes delicate species such as midges, gnats, and mosquitoes, and the Brachycera includes the heavier-bodied horse flies, deer flies, fruit flies, house flies, blow flies, bat flies, and so on. Flies undergo holometabolous development, and fly larvae (maggots) do not resemble the adults into which they mature. Most flies are oviparous; that is, eggs are laid that eventually hatch into larvae. Common oviparous flies include mosquitoes, gnats, sand flies, fruit flies, house flies, blow flies, flesh flies, most bot flies, horse flies, and deer flies. Some flies, including the flesh flies (Family Sarcophagi- dae), are ovoviviparous and larviparous; the eggs of flesh flies and some bot flies (including Cephenemyia spp., the nasal bots of deer) hatch inside the female's body and larvae are deposited directly onto the host (including mammals, birds, or other insects) or carcass (depending on the species). Still other flies, including the louse flies or keds (Family Hip- poboscidae) and the infamous tsetses of Africa (Family Glossinidae) are pupiparous; that is, eggs hatch inside tsetses and keds, and the larvae are retained inside the female until they are ready to pupate. Fully mature larval tsetses and keds are “born” immediately prior to their pupation in the environment. The considerable diversity of larval and adult forms of flies corresponds to patterns of natural history common within the phylogenetic groups (Hall and Gerhardt 2009). The life history characteristics and propensity to cause disease will be discussed for selected flies representative of the groups most commonly associated with wildlife diseases.
mosquitoes The 3,500 species of mosquitoes (Family Culicidae) reach their greatest diversity and densities in tropical forests, though temperate forests and Arctic tundra also support high densities during specific seasons. Mosquitoes transmit a greater diversity of disease causing agents (including viruses, bacteria, protozoa, and nematodes) than any other group of insects (Foster and Walker 2009).
Mosquito eggs are oviposited in aquatic habitats that range in size from tiny pools of water in animal or tire tracks, to pools of water associated with snowmelt, water sequestered in tree cavities, ephemeral ponds, permanent ponds, rivers, and streams, to large lakes; of course, specific characteristics of oviposition sites depend upon the biology of the species. Most mosquito larvae are filter-feeders, but larvae of some species are predators of other mosquito larvae. Wigglers float along in the surface tension of the water on clusters of long setae, and they undergo four molts, through four instars of increasing size, prior to pupation. Mosquito pupae are called “tumblers” because the pupae remain active, causing the puparium to tumble along beneath the water surface. Once mature, the puparium splits to release the adult (imago) mosquito. It is noteworthy that only female mosquitoes feed on blood which provides nourishment necessary for egg production in many species; species in which females do not need such a blood meal are termed autogenous (meaning self-generating) (Foster and Walker 2009, Hall and Gerhardt 2009).
The two subfamilies of the Culicidae, the Anophelinae and the Culicinae, differ in morphology, behavior, host preference, and ability to serve as vectors of different agents. The Anophelinae are represented by the genus Anopheles, which lay eggs singly into standing water; each egg has specialized floats to keep it on the surface. The Culicinae are represented by the common genera Culex and Aedes. Culex spp. oviposit into water as rafts of up to 400 eggs, while Aedes spp. oviposit in moist soil or in areas likely to be flooded (as by snow melt or in tree hollows likely to collect water). Whereas the eggs of Culex spp. and Anopheles spp. tend to hatch within about 5 days, the eggs of some Aedes spp. can survive years without desiccating, to hatch as the environment becomes saturated with water. Anoph- eline larvae lack breathing tubes, whereas culicine larvae have long breathing tubes that function as “snorkels.” Anopheles spp. adults can be identified from a distance because they tend to rest with their abdomens raised at an angle to the substrate on which they sit, whereas Aedes spp. and Culex spp. tend to rest with their abdomens in a plane horizontal to the substrate. Adults of both sexes of Anopheles spp. have long maxillary palps next to their oral proboscis, but female culicines have short palps.
The many important disease agents transmitted by mosquitoes include those causing malaria (Plasmodium spp.) of birds, rodents, and primates; West Nile fever virus; Rift Valley fever virus; avian pox viruses; myxoma virus; and filarial nematode worms (including several species of heart worms such as Dirofilaria imm- itus of canids, D. subdermata of porcupines, and D. magnilarvatum of macaques) (Allan 2001a, Foster and Walker 2009).
black flies (Family Simuliidae) are adapted to life in and around fast-moving streams, rivers, and melting glaciers. Females oviposit onto the surface of rocks or other substrate beneath the water surface. Once the larvae hatch, each spins a silken cord that serves as a lifeline anchoring it to the substrate on which it hatched. This arrangement results in a swarm of small, tethered, black fly larvae that filter-feed on small organic debris being swept along by the water. The larvae feed and molt within the current, eventually developing into adults inside anchored cocoons; in many species of black flies, the pupa is resistant to desiccation and can survive in habitats where water levels drop prior to emergence. Eventually, adults emerge from pupae and crawl out of the water to dry off, obtain meals, and produce more offspring (Allan 2001a, Adler and McCreadie 2009).
Many of the 2,000 species of black flies are important parasites of wildlife. Large swarms of these flies are associated with death from exsanguination of animals as large as domestic cattle (Allan 2001a). Moreover, black flies are important vectors of several parasites of wildlife, including blood protozoa of birds (Leu- cocytozoon spp.) and filarial nematodes (Onchocerca spp.) of ungulates and humans; they are also a source of considerable annoyance for animals in Arctic regions (Allan 2001a, Adler and McCreadie 2009).
sand flies are small, delicate flies in the Family Psychodidae and Subfamily Phleboto- minae, hence the commonly used name of phlebotomine sand flies. These small flies breed in dark, humid microhabitats containing abundant organic matter, including tree hollows, forest litter, cracks in soil or rocks, rodent burrows, cesspools, poultry and livestock barns, and so on. Larval sand flies feed on decaying organic matter and fungi, and it is the adult flies that serve as vectors of disease agents. The delicate adults generally lack the potential for sustained, long-distance flight and are typically found resting in areas near the breeding sites. Adult sand flies feed primarily on plant juices and sugars, but females of most species also feed on blood to provide the nutrition necessary for egg production. Some species of sand flies, such as Lutzomyia gomezi, feed on a broad range of birds and mammals, while others feed specifically on certain types of vertebrates; L. vexator feeds exclusively on lizards and transmits lizard malaria, while L. Vespertilionis feeds exclusively on bats (Allan 2001a, Rutledge and Gupta 2009).
Sand flies also transmit vesicular stomatitis virus to ungulates, opossums, primates, and some mammalian carnivores; Plasmodium mexicanum (causing lizard malaria); Leishma- nia spp. (causing leishmaniasis); and some species of Bartonella, including the zoonotic Bartonella bacilliformis (Allan 2001a, Rutledge and Gupta 2009).
midges of the Family Ceratopogonidae are flies whose bites cause a burning pain more severe than suggested by their tiny size. Because of the pain of their bites, people know them by a variety of common names including midges, gnats, no-see-ums, punkies, moose flies, and (although it tends to confuse students studying the group), in some areas they are referred to simply as flies. In addition, the salivary proteins responsible for the painful bites often cause considerable inflammation around the bite site, and many species of livestock (and presumably related mammals) develop severe allergic reactions to these flies. It is common for midge larvae to become dormant during either hot summer months or cold winter months, depending on the biology of the species. Such dormancy allows mass eruption of local populations when environmental conditions become right for emergence and breeding. Activity of most Culicoides spp. is seasonal and may be specific to time of day; some species are quite active during the heat of warm summer afternoons, but this varies considerably among species, and some species remain active year round. Some species are univoltine, producing only a single generation per year, associated with a specific season such as early spring, late spring, or summer, while others are multivoltine, producing several generations during a single season. As autumn progresses, the first period of cold weather is usually enough to drive populations into dormancy to await another summer season. In the United States, the genus Culi- coides represents the midges most often associated with wildlife diseases (including the reoviruses causing blue tongue and epizootic hemorrhagic disease of deer, bunyaviruses, rhabdoviruses, at least seven species of avian blood parasites of the genus Haemoproteus, and a number of filarial nematodes) (Allan 2001a, Mullen 2009).
HORSE FLIES AND DEER FLIES Unlike the delicate nematocerans discussed above, the horse flies and deer flies (Suborder Brachycera; Family Tabanidae) are large (6 to 30 mm), robust flies with strong wings and large, slashing mouthparts. Horse flies (Tabanus spp. and Hybomitra spp.) and deer flies (Chrysops spp.) are active during the day when they interact with people and other diurnal species. Tabanids lay egg masses on vegetation above wet substrates such as shrubs at the edge of a pond or stream. Larvae hatch and drop into the mud of aquatic or semiaquatic habitats, where they feed on organic debris, microorganisms, or other invertebrates (depending on the species, and some Tabanus spp. larvae are cannibalistic) (Allan 2001a, Mullen 2009). Predacious larvae (including Tabanus spp.) have mandibles modified into large, downward-projecting fangs that are used to capture and subdue their prey. Adult tabanids often lie in wait for potential hosts, but they are also strong fliers capable of regular flights of several kilometers in search of hosts on which to feed. The large slashing mouthparts of adult tabanid flies involve the paired maxillae and mandibles that slash “scissor-like” through the skin, creating deep wounds. Anticoagulants in the saliva are then pumped into the wound, ensuring that it bleeds profusely, and the blood is sponged up through the fly's labella. As with many other blood-feeding flies, taba- nids often bite within specific regions of the host's surface. Thus, some species may prefer to bite the legs, while others tend to bite along the belly or the back. These are also persistent feeders that will readily fly to a different host if their meal is interrupted, and such persistence has been shown to be important in the mechanical transmission (discussed below) of parasitic organisms. Tabanids are important mechanical vectors of the agents causing anthrax and tularemia, as well as the protozoan Besnoitia besnoiti, and they serve as the obligatory biological vector of Elaeophora schneideri, an arterial worm of cervids (Allan 2001a, Mullen 2009).
tsetses (Family Glossinidae) are endemic to sub-Saharan Africa and are often cited as an important factor that limited expansion of European colonists and their European breeds of cattle on the continent during previous centuries (Krinsky 2009b). Tsetses transmit the blood-borne protozoan parasites (Trypanosoma spp.) that cause African sleeping sickness in people and n'gana (or nagana) in cattle. They also transmit these, or similar, parasites among wild ungulates in areas where they are common. Tsetse life history is similar to that of muscid flies, except that only a single egg is produced during each gonotropic cycle, and it is fertilized by sperm stored from a copulation prior to ovulation. The egg hatches within the female's body and the larva is nourished by the female through the three molts of the larva. Females then larviposit the fully mature larvae onto the soil, into which it burrows and then pupates almost immediately. Thus, tsetses are extremely “K-selected” for insects in that their reproductive rate is very low and the female must invest heavily in each larva produced. Both male and female tsetses rely on vertebrate blood as their sole source of nourishment. Each of the 23 species of tsetses has its own habitat requirements, and host preferences vary considerably; one group of species feeds most commonly on reptiles (including crocodiles) and then opportunistically feeds on small mammals, ungulates, and humans; other species feed in open savanna or forested habitats mostly on ungulates, but some species also readily bite ostriches and small mammals (Krinsky 2009b). Thus, transmission of trypanosomes among wildlife, as well as to cattle and people, is restricted to specific habitats in which specific species of tsetses overlap with the distribution of reservoir hosts.
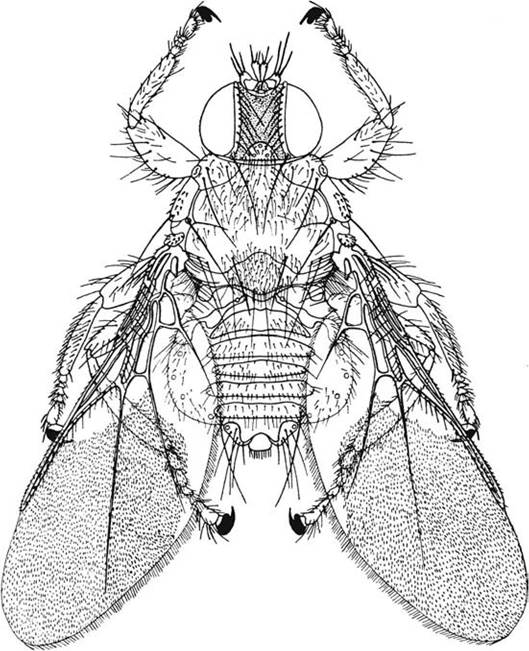
FIGURE 5.4 Ornithoica vicina is a bird-infesting ked (Family Hippoboscidae). These flies are dorsoventrally flattened and may be found with or without wings (depending on the species) while infesting vertebrate hosts (from Capinera 2010, copyright © John Wiley and Sons, by permission).
Hippoboscid flies Also called keds or louse flies, hippoboscid flies (Fig. 5.4) are common parasites found on birds or mammals (although no species is found on both birds and mammals) throughout the temperate and tropical regions. These dorso-ventrally flattened flies are commonly mistaken for lice or ticks, hence the name louse flies, but the forewings present on most species clearly separate them from these wingless ectoparasites. Readers familiar with ectoparasites of deer in North America may recognize the dorso-ventrally flattened keds of the genera Lipoptena and Neoli- poptena; these two genera are interesting in that the newly emerged adults are winged, but they shed their wings once they have successfully dispersed to a new host. Host specificity varies considerably among species, with some specialized on single host species and others feeding broadly across avian or mammalian taxa (Harwood and James 1979, Gullan and Cranston 2005, Lloyd 2009).
All hippoboscid flies are parasitic and adapted to obligatory blood-vessel feeding (Lloyd 2009). Like the tsetses described above, females larviposit a fully mature larva that pupates immediately. In some species, the puparium glues itself to the host's hair; in others, such as the pigeon fly, Pseudolyncha canariensis, larvae are deposited in the nesting material where they pupate.
Most hippoboscid flies cause relatively few problems for wildlife. However, they were recently associated with severe hair loss, poor body condition, and deaths of moose in Scandinavia (Madslien et al. 2011). Perhaps more importantly, keds are vectors of a number of species of the genus Haemoproteus (avian blood parasites), Dipetalonema dracuniculoides (Lloyd 2009), and have recently been implicated as potential vectors of Bartonella spp. (Halos et al. 2004). They are also thought to be important phoretic hosts, dispersing bird lice and feather mites (Jovani et al. 2001, Proctor and Jones 2004).
bat flies Bat flies in the Families Strebli- dae and Nycteribiidae are obligate blood-feeding parasites (Lloyd 2009). Host specificity varies considerably among bat flies, but many species, including those in the Americas, tend to be quite host specific. Moreover, many species of bat flies have adapted to survival on specific body regions, with different species restricted to the wing membranes, head, or body of their hosts (ter Hofstede et al. 2004, Lloyd 2009). The density of bat flies found on different species of Neotropical bats has been correlated to preferences of cavities or foliage for roost sites (ter Hofstede and Fenton 2005), but it may be difficult to determine cause and effect relationships. Although there is a growing literature on the occurrence of bat flies on bats (Chilton et al. 2000), there is little known about their potential roles as vectors of pathogenic agents among bats. Bartonella spp. are bacteria that infect red blood cells ofvertebrates, and several new genotypes were recently reported from bat flies; but the disease associated with these flies remains debatable (Morse et al. 2012).
filth flies, bot flies, flesh flies, and heel flies Flies in the Family Muscidae are commonly referred to as filth flies. These are quintessential flies for most people, and it is this group that is often thought of first when someone mentions the word “fly.” This family includes the house fly, Musca domestica, recognizable as robust, medium-sized, gray flies about 5 mm long with dark stripes along their thoraxes. Muscid flies undergo a life cycle similar to the most Diptera. They breed, oviposit eggs which hatch into larvae, undergo three molts, form pupae, and develop into adults that repeat the cycle. (Fig. 5.5). Adult filth flies typically lay eggs in organic environments that are moist enough to promote aerobic microbial fermentation, such as rotting vegetation, garbage, or animal fecal material (Moon 2009). In the case of filth flies (and related species of the Families Calliphoridae, Sarcophagidae, Oestridae, and some others), the larvae are commonly called maggots. Maggot bodies are tapered, and lack a sclerotized head capsule. Most obtain nourishment by filter-feeding on the substrate on which the eggs or larvae were laid. Feeding is accomplished by probing the substrate with hooks positioned near a ventral sieve, which serves as the mouth, at the pointed end of the larva. The anus and the respiratory spiracles are found terminally at the larger, posterior, end of the maggot. The characteristics of the mouth hooks and the respiratory spiracles are important for specific identification of maggots (Moon 2009).
Many muscid flies, including the house fly, have mouthparts adapted to sponging sugars or fluids from organic debris, rotting vegetation, or feces. However, some muscids have adapted their once-spongy proboscis into an elongated, hardened feeding tube that pierces the skin and through which blood and fluids are sucked during feeding; these muscids are commonly referred to as biting flies; they include the stable fly, Stomoxys calcitrans, and the horn fly, Haematobia irritans. Biting flies often prefer to feed on specific body regions; hence the name horn flies, indicating that this species feeds near the ears and head of cattle, bison, and other ungulates. Non-biting filth flies may serve as mechanical vectors of enteric pathogens (E. coli, Salmonella spp., Camplylobacter spp., etc.) picked up from manure or rotting organic matter that is then inadvertently carried to food or mucous membranes (including the eyes, mouth, teats, etc.). In addition, some species serve as intermediate hosts of internal parasites; the eye worm, Thelazia californiensis, is biologically transmitted among rabbits and other mammals in the western United States by the face fly, Fannia thelaziae. Although it would seem
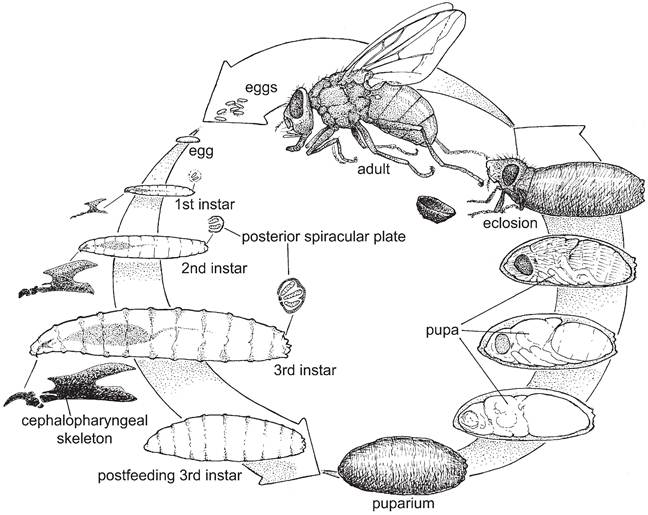
FIGURE 5.5 (A) Generalized life cycle of muscid and oestrid flies showing eggs, first, second, and third instar larvae (commonly called maggots), development of the pupa, and adults (from Mullen and Catts 2002, copyright © Elsevier, by permission).
that the blood-sucking biting flies have more intimate contact with the host and therefore provide more opportunities for transmission of blood-borne pathogens, biting muscids are associated with relatively few pathogens causing wildlife diseases (Moon 2009).
The blow flies (Family Calliphoridae), flesh flies (Family Sarcophagidae), and bot and warble flies (Family Oestridae) are similar in basic body shape to muscids, but they typically lay their eggs (or sometimes larvae) on or in the vicinity of live or dead animals. Larvae that hatch out progress through three molts while feeding on the tissues of animals (or carcasses) upon which the eggs were laid. These families were covered below under the subheading Myiasis.
More on the topic THE INSECTS:
- Other Insects
- FIVE The Parasitic Insects, Mites, and Tick
- Germs, parasites, insects and nematodes
- EXAMPLES OF DISEASES CAUSED BY INSECTS, MITES, AND TICKS
- CONTENTS
- THE MITES AND TICKS
- If you looked across a landscape from the top of a mountain, you would see a patchwork of different communities—say, forests, meadows, lakes, streams, and marshes (FIGURE 19.3).
- Insect Bite Dermatitis
- Introduction
- GENERAL INTRODUCTION