VECTOR-BORNE BACTERIA
As discussed previously, bacteria can be transmitted by arthropods mechanically (e.g., moved between hosts by simple contamination, without replication or development) or biologically (e.g., efficiency of transmission generally requires a period for multiplication of numbers in the vector).
Unlike some protozoa, fungi, and helminths that undergo necessary transitions through progressive life stages, bacteria do not undergo changes in life stage and, therefore, are not transmitted via developmental or cyclode- velopmental transmission cycles. This section includes descriptions of tularemia, plague, and Lyme disease as representative diseases caused by vector-borne bacteria with wildlife reservoirs.Francisella tularensis
causative agent Francisella tularensis is a gram-negative, aerobic, non-spore-forming rod (App. 1: Table 7), with at least four subspecies, Francisella tularensis biovar tularensis (=nearc- tica, also called Type A), F. tularensis biovar palearctica (=holarctica, Type B), F. tularensis mediasiatica, and F. tularensis novicida (Morner and Addison 2001, Morner 2007). This organism is a highly infectious bacterium capable of invading most tissues in its vertebrate and arthropod hosts (Sonenshine 1993, Ellis et al. 2002, Dennis 2004), and this high level of infectivity for different types of tissues accounts for its range of potential hosts and transmission routes (Brown et al. 2005, Foley and Nieto 2010, Telford and Goethert 2010).
HOST RANGE AND GEOGRAPHIC distribution Francisella tularensis originally was assigned its Latin name because it was identified from Tulare County, California during surveys for plague in ground squirrels (Spermophilus beecheyi) (Hopla and Hopla 1974, Morner and Addison 2001). However, as often occurs with species named for their places of discovery, the organism occurs over a much broader geographic range (from the Arctic to subtropical regions) than the name would suggest (Brown et al.
2005). The two classic pathogenic subspecies are F. tularensis tularen- sis (type A strains, known mainly from North America) and F. tularensis holarctica (Type B strains, found throughout the Holarctic region) (Morner and Addison 2001). These strains vary in their patterns of ecological maintenance and in their virulence for hosts; F. tularensis tularensis (type A) is the more virulent form (Dennis 2004, Foley and Nieto 2010, Telford and Goethert 2010).This organism occurs only in the Northern Hemisphere and has one of the broadest host ranges of any bacterium, with reports from 190 species of mammals, 26 species of birds, three species of amphibians, and 88 species of invertebrates (Morner and Addison 2001, Morner 2007). However, these bacteria primarily parasitize rodents and lagomorphs (Hopla and Hopla 1994), and may have co-evolved with these groups in the Northern Hemisphere (Olsufiev 1959). Birds do not play an important role in the epidemiology of tularemia (Morner 2007).
RESERVOIRS AND TRANSMISSION Lago- morphs and rodents serve as reservoir hosts for F. tularensis. In North America, rabbits (Sylvila- gus spp.), hares (Lepus spp.), muskrats (Ondatra zibethica), beavers (Castor canadensis), voles (Microtus spp.), and squirrels (Sciurus and Spermophilus spp.) have been implicated as reservoir hosts (Acha and Szyfres 2001, Morner and Addison 2001, Dennis 2004). Most rodents and rabbits are susceptible to acute systemic infections. However, some small mammals, including the European rabbit (Oryctolagus cuniculus), and carnivores are less susceptible or resistant to development of disease (Morner and Addison 2001, Dennis 2004). Historically, many human cases in North America were acquired by hunters of cottontail rabbits; hence the common name “rabbit fever.” However, even in North America, the ecological importance of rabbits as reservoir hosts may vary considerably among areas (Telford and Goethert 2010).
Francisella tularensis grows well in water and mud, and infectious rodents contaminate environmental sites, amplifying the epizootics beyond the reservoir as new species become exposed at water sources (Parker et al.
1951, Tarnvik et al. 2004). Water-borne transmission is the common denominator in epizootics in central Europe and Asia involving F. tularensis holarctica (Type B) (Morner 1992, Tarnvik et al. 2004, Foley and Nieto 2010). In typical epizootics, beavers (Castor spp.), muskrats (Ondatra zibethicus), and water rats (Arvicola terrestris) promote the transmission cycle. As an interesting aside, this pathogen’s profound ability to infect a variety of hosts and vectors has provided the traits exploited for use as a biological weapon (Dennis et al. 2001, Chang et al. 2003).The primary routes of transmission include arthropod bites (including those of ticks, taba- nid flies, mosquitoes, and black flies), mechanical transmission by arthropods, contact with contaminated water, contact with tissues of infected animals, or even by aerosol transmission. Because the role of arthropod transmission is so important, we consider this bacterium as an arthropod-borne parasite.
While many arthropods can serve as mechanical vectors, several species of ticks are competent biological vectors of F. tularensis, and ticks may serve as an important interepizootic reservoir of infection (Acha and Szyfres 2001, Morner and Addison 2001). The rabbit tick, Haemaphysalis Ieporispalustris, specializes on rabbits throughout the Holarctic region and is considered an important enzootic vector of F. tularensis in many areas. Ticktransmission to other mammals (including humans) is typically associated with bridging vectors, including Dermacentor spp. and Amblyomma americanum in North America, and Ixodes apronophorus and Dermacentor retic- ulatus in Europe and Asia (Sonenshine 1994, Hubalek et al. 1996, Brown et al. 2005).
There are three ecological patterns of zoonotic transmission in northern Asia (Morner and Addison 2001). Humans in North Asia most commonly contract tularemia during spring, in association with epizootics in water rats, Arvicola terrestris, and exposure occurs either from direct contact with dead or dying animals or via infected water sources.
A second type of transmission is common in this region during the autumn months, when exposures result from high densities of mosquito vectors. Third, people become exposed while hunting or trapping rabbits during the winter months; thus, transmission routes can be complicated even when one route is thought to predominate.In northern Europe and Scandinavia, the most common human exposures occur during years coincident with epizootics of varying hares and are associated with mosquito bite rates, which peak during late summer and early autumn (Morner and Addison 2001, Dennis 2004, Brown et al. 2005). In arid regions of the western United States, exposures to F. tularensis tularensis (Type A) are most common during summer months when deer flies (Chrysops spp.) and horse flies (Tabanus spp.) serve as mechanical vectors; seasonality of exposures is closely tied to vector activity in this region (Krinsky 1976, Morner 1992, Morner and Addison 2001, Petersen and Schriefer 2005). However, even relatively close areas, such as Martha's Vineyard and Nantucket Island in Massachusetts, may differ significantly in the ecological maintenance as well as transmission risks to humans; one obvious difference noted between the islands was the larger populations of American dog ticks on Martha's Vineyard where hosts (various wild carnivores and larger mammals) are abundant (Telford and Goethert 2010).
clinical effects In susceptible host species, including humans, the disease typically begins with local areas of tissue necrosis associated with multiplication of organisms at the site of colonization. Bacteria entering and multiplying in the blood vascular system cause bacteremia and septicemia that spread the infectious agent throughout the viscera. The resulting colonization creates pale, necrotic foci throughout the liver, spleen, and bone marrow (Morner and Addison 2001, Dennis
2004). Hunters and trappers of rabbits, muskrats, and beavers may recognize the characteristic white spots on the livers of infected animals, suggestive of tularemia.
In addition, lungs may become congested and edematous, with regions of consolidation associated with pneumonia and pleuritis; lymph nodes may have foci of caseous necrosis, and granulomatous lesions may be evident in the kidneys, liver, lungs, or spleen (Morner and Addison 2001, Foley and Nieto 2010). Sick animals may appear apathetic and may die of septicemia during the first two weeks following infection. In somewhat less sensitive species, the progression of the disease may be more chronic, with signs including ulceration of the site of infection, fever, swollen lymph nodes, depression, pneumonia, and chronic wasting (Morner and Addison 2001, Foley and Nieto 2010).population effects Most epizootics of tularemia are sporadic, and epizootics among small mammals often go undocumented. However, mortalities among rodent fur-bearers (muskrats and beavers) and lagomorphs hunted for sport have been associated with tularemia (Morner and Addison 2001, Telford and Goethert 2010). To a large degree, the impacts of an epizootic of tularemia depend on the rate of transmission among hosts, and highest rates of transmission occur when vector densities are high or when populations of aquatic rodents are high (Morner and Addison 2001, Dennis 2004). Management problems may arise when large numbers of a preferred species (fur-bearers, cottontail rabbits, etc.) die or when the habitat is altered as a result of changes in the community structure of primary consumers (Morner and Addison 2001). The greatest problems for conservation are expected when dense populations of reservoir hosts are sympatric with small and isolated populations of susceptible rodents or lagomorphs (Cleaveland et al. 2002).
special problems No special problems currently are identified.
control Control of F. tularensis is mainly limited to public education about the local risks of exposure to reduce the number of human cases. Thus, recognition of the common routes of transmission, seasonality of local vector activities, methods for avoiding biting vectors, and signs of disease in small game mammals is essential.
YERSINIA PESTIS
causative agent The term plague can be used generally to refer to any pestilence or devastating epizootic, but, more specifically, it refers to the disease caused by Yersinia pestis. Yersinia pestis is a small, pleomorphic, facultatively anaerobic, gram-negative bacillus (Family Enterobacteriaceae; App. 1: Table 7) that is widely distributed through tropical and temperate regions (Acha and Szyfres 2001, Abbott and Rocke 2012).
host range and geographic distribution Plague is thought to have evolved on the steppes of Asia; it has been responsible for over 200 million human deaths during three global pandemics recorded in history; half of those occurred during the Justinian Plague, which began in the sixth century (Acha and Szyfres 2001, Gasper and Watson 2001, Eisen and Gage 2012). Indeed, the current geographic distribution of plague in the world is a patchy relict of the most recent pandemic, which began in China in the 1850s (Gasper and Watson 2001, Gage and Kosoy 2005, Eisen and Gage 2012). Contemporary plague is commonly reported from Africa, Asia, and South America, and there were 999 cases reported in the United States between 1900 and 2010 (Centers for Disease Control 2012). Although the ecology of Y. pestis varies from region to region, and involves a large number of different hosts and vectors, some general ecological patterns seem consistent among the regions (Gasper and Watson 2001, Gage and Kosoy
2005). Comparison of host distributions and that of plague in North America has shown that the geographic distribution of plague is more related to niche parameters (and the trophic cascade hypothesis), including temperatures related to latitude and elevation, precipitation, and flea species distributions, rather than the distribution of specific reservoir hosts (Maher et al. 2010, Eisen and Gage 2012).
reservoirs and transmission Ecological maintenance cycles often are complicated and vary among biomes and geographic regions because of differences in the abundance and distribution of vectors and reservoir hosts. Wild, feral, and peridomestic rodents serve as reservoirs of Y. pestis, and the agent is transmitted by rodent-feeding fleas in all regions where plague persists enzootically. In addition, there are epidemic cycles involving human populations (sometimes referred to as urban plague) that occur only when human densities are high, hygiene is compromised, and circumstances facilitate human-to-human transmission (Acha and Szyfres 2001, Gasper and Watson 2001, Gage and Kosoy 2005, Abbott and Rocke 2012). Urban plague is further supported by epizootic transmission among peridomestic rats (Rattus spp.) and people that contact infective rat fleas, including the cosmopolitan Oriental rat flea, Xenopsylla cheopis. Although human plague is an interesting subject, we will focus the remainder of our discussion of sylvatic plague and the maintenance of enzootic cycles and disease in wildlife.
More than 200 species of rodents, and 14 species of lagomorphs, have been found infected with Y. pestis (Durden and Traub 2002), but, due to variability in host resistance and susceptibility, only a small percentage of these serve as stable reservoirs of plague (Gage and Kosoy 2005, Abbott and Rocke 2012). Ecological maintenance in stable transmission cycles typically is supported by a few species of rodents in which infections persist long enough to facilitate transmission and of which some may survive the infection. Reservoir hosts often are fecund, with multiple generations per year, ensuring a regular supply of susceptible hosts, and tend to have short life spans (Gage and Kosoy 2005, Abbott and Rocke 2012).
In North America, voles (Microtus spp.), deer mice (Peromyscus spp.), grasshopper mice (Onychomys spp.), and kangaroo rats (Dipodo- mys spp.) appear to be somewhat resistant; in Asia, resistance has been reported in marmots (Marmota spp.) and gerbils (Rhombomys opimus and Meriones spp.) (Gasper and Watson 2001, Gage and Kosoy 2005). Individuals within populations of several species (gerbils, marmots, deer mice, and California voles) vary in their resistance; some individuals suffer severe disease and die, while others survive after mild infections (Gage and Kosoy 2005, Abbott and Rocke 2012). Heterogeneous resistance also has been demonstrated among populations of grasshopper mice, deer mice, California voles, and prairie dogs that vary in their historical associations with Y. pestis (Gasper and Watson 2001, Gage and Kosoy 2005); changes in gene frequencies that promote resistance have been reported from some populations that have suffered repeated epizootics of plague.
Other rodent species typically develop high levels of bacteremia, die quickly, and may serve to amplify infections beyond enzootic foci during epizootics. Within the United States, prairie dogs (Cynomys spp.), ground squirrels (Otospermophilus spp.), and wood rats (Neotoma spp.) often serve as amplifying hosts, and nearly 100% of infected animals may die during epizootics (Ubico et al. 1988, Gasper and Watson 2001, Trudeau et al. 2004, Gage and Kosoy 2005). Amplifying hosts are important because they infect large numbers of fleas, spread the pathogen to additional species of vectors, move the pathogen beyond the enzootic focus (nidus), and generally enhance the probability of a rapid increase in the number of infected hosts and vectors.
Susceptibility varies considerably among the orders of mammals; primates, some carnivores (felids and some mustelids), lagomorphs, and most species of rodents tend to be highly susceptible, and often die, whereas other carnivores (canids and ursids), ungulates, and some rodent species appear to be fairly resistant to Y. pestis; it is important to note that domestic cats and black-footed ferrets (Mustela nigripes) suffer high probabilities of mortality when infected with Y. pestis (Williams et al. 19 91, 1994; Gasper and Watson 2001; Gage and Kosoy 2005; Abbott and Rocke 2012). Thus, vertebrate hosts have often been classified as one of four types of species, each serving different roles in the epizootiology of plague: (ι) rodent species that serve as the enzootic reservoir; (2) rodent species that amplify infections beyond enzootic foci; (3) resistant non-reservoir hosts, birds, ungulates, and most carnivores, that are generally unimportant to plague transmission; and (4) susceptible non-rodent hosts (humans, some mustelids, cats, and some rodents) that may promote some transmission but which generally are not involved in the ecological maintenance (Gasper and Watson 2001) (Fig. 9.1).
Yersinia pestis typically is transmitted by the bite of vector-competent fleas. More than 1,500 flea species have been associated with Y. pestis after having become infected while feeding on blood and lymph f luids of an infected host (Gage and Kosoy 2005), and approximately 250 flea species have been reported as naturally infected (Eisen and Gage 2012). However, flea species differ dramatically in their ability to serve as vectors in cycles and in the accidental transmission to aberrant hosts, and most of these species are probably
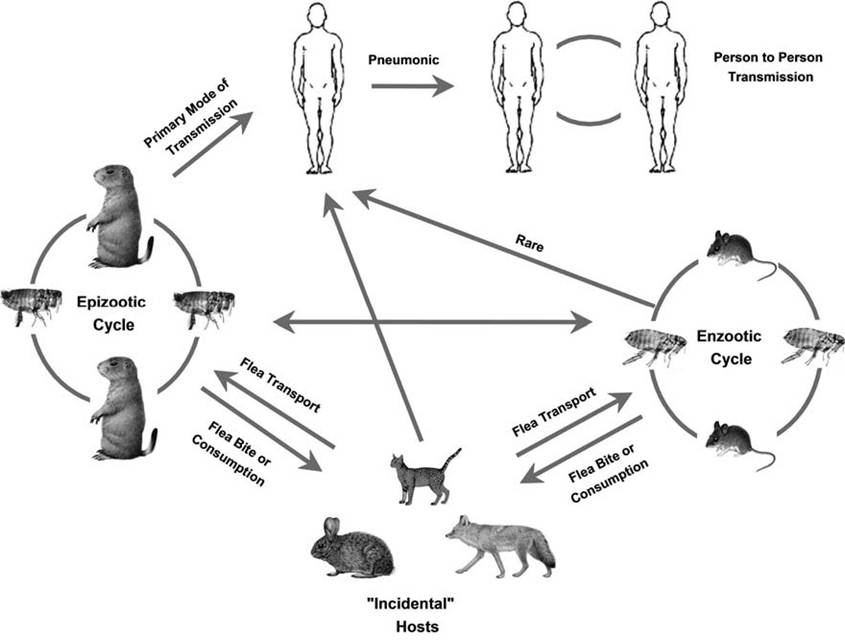
FIGURE 9.1 A simplified model of natural cycles of plague. Sylvatic cycles are maintained by fleas and wild rodents in North America. Enzootic cycles maintain the pathogen in nature. When conditions support spillover into non-typical reservoir hosts, then epizootics may occur. Incidental hosts become infected, some of which may suffer severe disease, and they may move the pathogen to new communities of potential amplifying hosts which support epizootic cycles. Specific species of reservoir hosts and fleas vary among different regions in the world and wild species commonly involved in cycles in the United States are illustrated here; note that risk of human to human transmission is expected to be very low in North America, but is much greater in underdeveloped regions where peridomestic rats and their fleas are uncontrolled and where human population densities are high (from Gage and Kosoy 2005, copyright © Annual Reviews, Inc., by permission).
inefficient or accidental vectors. Internal morphology and gut physiology are generally considered to be key aspects determining vector competence of different flea species.
Vector competence is defined as the tendency of a vector species to become infected, remain infectious, and then transmit to naive hosts. Vector competence of fleas for Y. pestis varies with ambient temperature; temperatures ≥28°C may reduce efficiency of transmission by X. cheopis due to enzymatic breakdown of the bacterial mat (Cavanaugh 1971, Khalil et al. 1984, Thomas et al. 1993). Thus, cooler temperatures that promote mat formation and retention may promote transmission, whereas higher temperatures reduce mat formation and facilitate clearance of the bacteria from fleas. Plague is not found in the hottest deserts, and this may result from a combination of inefficient transmission and desiccation and death of fleas when relative humidity is low (Gage and Kosoy, 2005). Rather, plague is typically associated with specific habitat types, including grassland steppes, cold high deserts, and mountain meadows. In arid regions, fleas often occur in burrows or nests that provide a microclimate with higher relative humidity than experienced in ambient environments. Even in the cooler desert regions in which Y. pestis can persist, there usually is less plague transmission during hot or drought years than in years with mild temperatures and higher humidity (Gage and Kosoy, 2005).
The Oriental rat flea is the best studied vector of urban plague because it feeds on a variety of rodents, including urban-dwelling rats, and due to its anthropophilic (human biting) habits (Gasper and Watson 2001, Ruiz 2001, Trudeau et al.
2004). Bacterial mats develop fairly quickly in X. cheopis (1-3 weeks), and the mats effectively block the gut. When a starving flea attempts to suck blood into its gut, the blood meal comes into contact with the bacterial mat. In addition, blocked fleas may repeatedly regurgitate their blood meals in attempts to dislodge the blockages; starving fleas may continue to attempt to feed and eventually die ifthey fail to dislodge the mat (Gasper and Watson 2001, Gage and Kosoy 2005). Whether the vectors die or survive may be of little consequence to the transmission ecology as long as Y. pestis is transmitted prior to flea death. However, many species of fleas transmit plague bacilli among rodent hosts without blocking, or in the early phase prior to development of the bacterial map. Transmission from unblocked X. cheopis may be as efficient as transmission via the classical mechanism (Eisen et al. 2007), illustrating that the traditional models of plague transmission may not be representative of all vectors and the actual list of important vectors may be much longer than recognized previously (Gage and Kosoy 2005; Eisen et al. 2007, 2008; Eisen and Gage 2012).
Whereas X. cheopis is an important vector in urban plague cycles involving Rattus spp. and potentially humans, most of the other 1,500+ species serve as vectors in sylvatic cycles involving wild rodents and their fleas. In the western United States, Oropsylla (Diamanus) spp. (including O. montana, O. hirsute, and O. tuber- culata), Hoplopsyllus anomalus, and Orchopeas spp. (including O. howardi and O. sexdentatus) are recognized as important sylvatic vectors (Gasper and Watson 2001, Gage and Kosoy
2005). Sylvatic vectors support local transmission cycles among wildlife and are of more importance to persistence of local and regional cycles of maintenance than is X. cheopis, but X. cheopis is clearly the most important urban vector on the global scale. Sylvatic rodent fleas may be found in large numbers in rodent burrows, and some retain their infections for prolonged periods following the deaths of their hosts. Infected fleas (O. tuberculata and O. labis) were recovered from prairie dog burrows more than a year following decimation of the prairie dog colony by plague (Kartman et al. 1962), and Gage and Kosoy (2005) discuss several studies of Asian fleas with proven ability to bridge periods of host inactivity. Thus, in addition to serving as a primary means of transmission of Y. pestis, some fleas provide a persistent reservoir.
As with the agent causing tularemia, Y. pestis is highly infectious and can be transmitted directly as well as by flea bite (Gasper and Watson 2001, Gage and Kosoy 2005). Yersinia pestis also is contagious and can be spread by ingestion or via respiratory aerosols, given the right set of circumstances (Acha and Szyfres 2001, Abbott and Rocke 2012). Carnivores and scavengers become exposed to infected prey during epizootics when direct contamination of wounds associated with feeding provides an important source of infection. Many predatory species, including bears (Ursus spp.), red foxes (Vulpes vulpes), coyotes (Canis latrans), and avian raptors tend to develop minimal disease when exposed, but they still may promote spread of the disease by moving infected carcasses or fleas. Additionally, most of these hosts develop antibodies following exposures (Gasper and Watson 2001, Gage and Kosoy 2005), and carnivores may provide good sentinel hosts heralding amplification of plague through rodent communities (Gasper and Watson 2001, Gage and Kosoy 2005, Abbott and Rocke 2012).
In North America, plague epizootics have caused 100% mortality of some prairie dog colonies (Trudeau et al. 2004, Biggins et al. 2010, Abbott and Rocke 2012). Such high levels of mortality have led to questions about the mechanism that maintains persistence of Y. pestis within prairie dog populations. One answer involves a combination of site fidelity by prairie dogs and alternative hosts, including grasshopper mice (Onychomys spp.); grasshopper mice become infected during epizootics and serve to promote persistence on the landscape by moving infected fleas between prairie dog family groups within large prairie dog colonies (Stapp et al. 2008, Salkeld et al. 2010). This model of transmission ecology is similar to a metapopulation model in which colonization of new prairie dog family groups by Y. pestis is dependent upon local abundance of sympatric grasshopper mice (Salkeld et al. 2010). Such findings suggest that the importance of alternative hosts, and multihost systems, needs to be reassessed in other regions of the world where plague is reportedly maintained by single-host reservoirs (Gage and Kosoy 2005, Salkeld et al. 2010).
clinical effects Three general classifications of disease are recognized in hosts that are susceptible to Y. pestis infections: bubonic plague, pneumonic plague, and septicemic plague. Bubonic plague involves bacilli localized in swollen lymph nodes and is typically transmitted by flea bite or predatory consumption. The term “bubonic” refers to the “bubos” that develop as grossly swollen lymph nodes in the axillae of infected humans and that are a common sign of Y. pestis infections. Pneumonic plague is transmitted via a respiratory route and tends to cause a rapidly fatal pneumonia. Septicemic plague involves bacteria that multiply in the blood stream, causing dissemination and shock that rapidly progresses to death. In humans and other susceptible hosts, pneumonic and septicemic plague cause mortality in approximately 100% of recognized, untreated cases (Gasper and Watson 2001, Gage and Kosoy 2005, Abbott and Rocke 2012). Although many carnivores are at least partially resistant to Y. pestis, domestic cats can develop clinical plague and, like humans, have a tendency to develop pneumonic plague. Unfortunately, this predilection of cats to develop pneumonic plague has led to numerous human cases resulting from respiratory inhalation of aerosol droplets from pets to their people (Gasper and Watson 2001, Gage and Kosoy 2005). Likewise, person-to-person transmission of pneumonic plague may occur during epidemics in crowded urban centers (Gasper and Watson 2001, Gage and Kosoy 2005, Abbott and Rocke 2012).
Wildlife biologists need to know the disease risks associated with wildlife with which they work, and they should wear appropriate personal protective equipment (PPE) during handling, sampling, and necropsies of wildlife. In 2007, an experienced, 27-year-old carnivore biologist died from plague approximately 1 week after necropsying a female cougar (Puma concolor). This cougar was part of a radio telemetry study when the biologist received a mortality signal from the transmitter. The biologist necropsied the carcass without gloves or other PPE (Wong et al. 2009). Many mammalian carnivores can be infected by Y. pestis, but felids are susceptible to developing pneumonic and septicemic plague (Abbott and Rocke 2012), both of which can be transmitted easily to a person who doesn't take adequate precautions during necropsy. In this case, knowledge of the disease risks associated with the animals being studied (and necropsied) might have saved this biologist's life.
population effects Epidemics of plague can decimate populations of susceptible rodents, rabbits and hares, and humans. Such decimation occurs occasionally at larger, widespread or regional spatial scales, during major epidemics, but more often it occurs seasonally or irregularly at smaller, local spatial scales. During periods between epidemics, Y. pestis may remain present in soil, in fleas, or associated with periods of inactivity or host hibernation (Gage and Kosoy 2005, Abbott and Rocke 2012). Like most diseases, the greatest potential impacts on populations are expected to occur when numbers are small and when immigration is limited.
special problems Dramatic reductions of prey populations also affect predator populations, especially when predators are specialists. For example, the endangered black-footed ferret (Johnson 2001, Biggins et al. 2010) depends upon prairie dogs (Cynomys ludovicianus and Cynomys leucurus); this small carnivore uses prairie dog burrows for cover and prairie dogs for prey. The species once ranged throughout the prairie grasslands of the United States, but was thought extinct in the wild in 1974 (Thorne and Williams 1988, Abbott and Rocke 2012). A surviving population was found in Wyoming in 1981, but concurrent epizootics of plague in the prairie dogs and ferrets, coupled with canine distemper virus in the ferrets, threatened to drive the population into extinction in the mid-1980s (Thorne and Williams 1988). The last remaining wild black-footed ferrets were captured in 1986 and 1987 to initiate a program of captive breeding and release. Although this program has been successful, prairie dog populations remain susceptible to local extinction with epizootics of plague, and the conservation of ferrets depends upon long-term persistence of prairie dog colonies (Dobson and Lyles 2000, Biggins et al. 2010, Abbott et al. 2012). This, of course, is only one example of population-level impacts; other examples could be found wherever plague is endemic.
control Unlike tularemia, risks of plague transmission are actively controlled in the western United States, as well as in many areas of northern Asia. Such control includes active surveillance for Y. pestis in known rodent reservoirs, control of domestic rats and fleas around houses or areas frequented by people and pets, control of wild rodents and fleas around houses or areas frequented by people, reduction in plague reservoir populations, and environmental management of sylvatic vectors focused on wild rodent burrows. Surveillance generally involves the reporting of sudden population declines in reservoir or amplification host populations. Rodents and associated fleas can be submitted to state or federal diagnostic laboratories equipped to safely isolate Y. pestis.
Active plague transmission is identified by a combination of confirmation of the organism in rodents during an epizootic, plague samples confirmed during surveillance, and confirmed human cases; once recognized, steps should be taken to alert the authorities and the public about local risks (Abbott and Rocke 2012). Further surveillance studies may be directed to determine which species of fleas are active in rodent burrows and the relative index of flea numbers (the flea index). Flea control can be achieved by direct application of insecticide dusts to burrows (when burrows are obvious, as with most ground squirrels, prairie dogs, etc.) or by setting baits that require feeding rodents to crawl through or over insecticide dusts (when burrows of rats and mice are too numerous or difficult to detect) (Barnes et al. 1974, Kimsey et al. 1985, Maupin et al. 1991, Kumar et al. 1997, Davis et al. 2002, Seery et al. 2003, Abbott and Rocke 2012). Campaigns to kill rodent hosts must be consistently applied and monitored to achieve effective plague control, and this method may be contraindicated once an epizootic is fully under way. However, the killing of hosts also can release large numbers of infective, host-seeking fleas, increasing local public health risks. Obviously, managers should carefully monitor any reduction of populations of conservation concern or that support populations of concern.
Vaccines can be used to protect highly susceptible hosts (including humans, prairie dogs, and black-footed ferrets) and to slow epizootic waves (Ruiz 2001, Creekmore et al. 2002, Abbott et al. 2012). Vaccine strategies used to control plague in wildlife depend on the ability to increase the ratio of resistant to susceptible hosts, and have generally depended upon injectable vaccines. For example, captive-bred black-footed ferrets have been vaccinated prior to release, and wild- born juveniles have been trapped and vaccinated. Raccoon poxvirus recently has been modified to express proteins of Y. pestis on its surface; although wildlife are not susceptible to disease caused by the recombinant virus, prairie dogs do develop immunity to the Y. pestis. Importantly, this new vaccine is being developed as an oral bait that seems to be highly efficacious (Abbott et al. 2012). Since many rodents (including the reservoirs of plague) are relatively short-lived and highly fecund, and population immunity depends on recent exposure to either the agent or the vaccine, the ratio of resistant to susceptible hosts would quickly revert to normal levels without continuation of regular (at least annual) administration of a vaccine; and effective vaccine delivery via oral baits provides hope for the conservation of prairie dogs, black-footed ferrets, and other wildlife species that use the burrow habitat created by prairie dogs (Abbott et al. 2012).
BORRELIA BURGDORFERI
causative agent Borrelia burgdorferi is a spirochete of the Family Spirochaetaceae (App. 1: Table 7) that causes Lyme borreliosis. Lyme borreliosis (Lyme disease) is the most commonly reported vector-borne disease of humans in North America (Bacon et al. 2008) and may result in as many as 255,000 worldwide cases annually (Rudenko et al. 2011). This is a tick- borne, zoonotic disease that results from infection by a group of related bacteria referred to as B. burgdorferi sensu lato (sl; meaning “in the broad sense”). This group of spirochetes includes over 18 named “genospecies” and an unknown number of “types” that remain to be classified (Bergstrom et al. 2002, Richter et al. 2004, Steere et al. 2005, Rudenko et al. 2011). Three main genospecies, Borrelia burgdorferi sensu stricto (ss; meaning strains that are genetically similar or “in the strict sense”), Borrelia afzelii, and B. garinii, commonly are associated with most human cases of Lyme disease (Steere 2001, Piesman 2002, Steere et al. 2005); in addition, Borrelia bissettii (Strle 1999), Borrelia spielmani (Richter et al.
2004), and Borrelia lusitaniae (Collares-Pereira et al. 2004) have been isolated from a small number of human patients in Europe. Although this description implies a considerable, and perhaps overwhelming, amount of diversity, this is a highly variable group of bacteria; one recent study reported 53 distinct genotypes within the narrow group referred to as B. burgdorferi ss (Crowder et al. 2010). The term “Lyme disease” is in common usage for human disease, but “Lyme borreliosis” has a broader meaning and is used to describe infections without necessarily implying that they cause pathology and disease; borreliosis is thus a more accurate term for infections in wildlife.
host range and geographic distribution Borrelia burgdorferi sl occurs mainly in temperate regions of the Holarctic (Asia, Europe, and North America) and extends into the subtropics of Southeast Asia (Eisen and Lane 2002, Brown et al. 2005). Many species of mammals, and some birds, become infected with B. burgdorferi sl. The range of potential hosts is broad, but specific groups of rodents or birds serve as reservoirs for each of the many genospecies of the spirochetes. Variation in nature is an axiom of biology, whether one is speaking of pathogens, vectors, or hosts. However, students of Lyme borreliosis must always consider which of the genotypes is being discussed and that general statements may be too broad to be accurate for any specific genotype.
reservoirs and transmission Borrelia burgdorferi sl is transmitted primarily by ticks in the Ixodes ricinus group, which includes several infamous disease vectors. The I. ricinus group includes the black-legged tick (also called the “deer tick”), Ixodes scapularis, in the eastern and upper-midwestern United States; the western black-legged tick, I. pacificus, in the far western United States; the taiga tick, I. persulcatus, in northern Asia; and the sheep tick, I. ricinus, in Europe (Eisen and Lane 2002, Brown et al.
2005). Each of these species feeds on a broad variety of vertebrate hosts, including rodents and birds, whose populations serve as reservoirs. Generally, larval and nymphal I. ricinus group ticks feed on small mammals, birds, and lizards, and adults feed on medium-sized or large mammals. Additionally, other Ixodes spp., including I. affinis, I. angustus, I. dentatus, I. hexagonus, I. japonica, I. jellisoni, I. minor, I. muris, and I. spini- palpis, may serve as vectors of B. burgdorferi sl.
When two or more cycles involving multiple vectors and borrelial strains co-occur in a host community, there are several possible effects. The cycles may be broadly mixed; in this case the two vectors both transmit two or more bor- relial strains. In other cases the cycles may be parallel, each vector transmitting its own strain among the same species of hosts. Or the cycles may be independent, each vector transmitting its own strain among different hosts (Eisen and Lane 2002, Gern and Humair 2002, Piesman 2002, Brown et al. 2005). Concurrence of cycles of borreliae in wildlife communities, involving both human pathogens and genospecies of
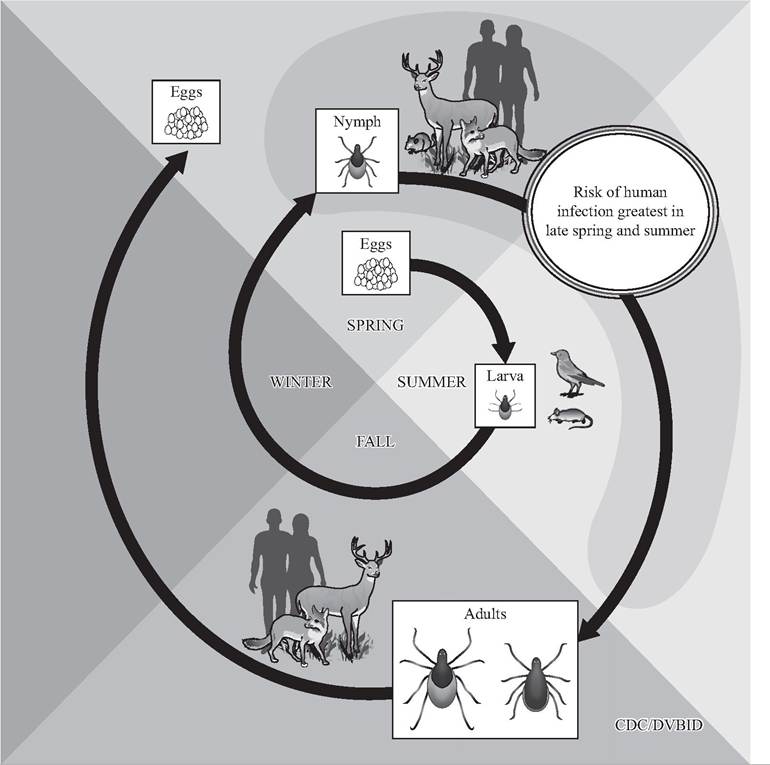
FIGURE 9.2 Generalized life cycle of Ixodes scapularis in the northeastern United States. The feeding of infected nymphs prior to name larvae amplifies the prevalence of Borrelia burgdorferi sensu lato in vector populations (Courtesy of Centers for Disease Control and Prevention, Division of Vector-Borne Diseases, http://www.cdc.gov/ncezid/dvbd).
unconfirmed pathogenicity, result in complex models of enzootic maintenance.
The generalized pattern for the transmission of B. burgdorferi sl in the northeastern United States involves naive (uninfected) larval ticks that acquire borreliae while feeding on infected rodent reservoir hosts (Fig. 9-2) (Spielman et al. 1985, Piesman 2002); transovarial transmission of B. burgdorferi sl from a female to her eggs occurs only rarely and is not considered an important part of ecological maintenance. Spirochetes survive the transstadial molt, multiply, and may disseminate within competent vectors. Some of the hosts fed on by infectious vectors will be naive to the infection, and the prevalence will increase in the host population. Infected nymphal and adult ticks transmit the agent beyond the enzootic cycle when they feed on susceptible vertebrates (Piesman 2002), but such hosts are often “accidental” or “dead-end” hosts that don't contribute to the ecological maintenance cycle.
Many small rodents are short-lived, with mean life expectancies of less than one year. Thus, infected ticks may live longer than many of the infected rodents, serving as a necessary bridge between seasons of tick activity. In the northeastern United States, prevalence in I. scapularis is promoted by the patterns of seasonal activity (feeding phenology) of the vectors. Importantly, the cohort of nymphs infected as larvae during the previous spring or summer feeds on white-footed mice (Pero- myscus leucopus) earlier in the spring than do the uninfected larvae. This reversed phenology results in naive larvae feeding on hosts recently infected by nymphs and amplifies the prevalence of infection in the vector population; this, in turn, greatly increases the probability of transmission to both reservoir and accidental hosts, including humans (Spielman et al. 1985, Piesman 2002).
The ticks that serve as the primary vectors of B. burgdorferi sl are generalist vectors that feed on a wide variety of vertebrate hosts, but most wildlife are incompetent as reservoir hosts (Ostfeld and Keesing 2000). Additionally, the hosts and vectors vary among different geographic regions and for the different genospecies of B. burgdorferi sl (Eisen and Lane 2002, Brown et al. 2005). For example, the primary reservoir of B. afzelii, a member of the B. burgdorferi sl complex in western Europe, includes the vole Clethrionomys glareolus, mice in the genus Apodemus, and tree squirrels (Sciuris spp.) (Craine et al. 1997, Humair and Gern 1998, Gern and Humair 2002). Avian reservoirs support other members of the complex, B. garinii and B. lusitaniae, in Europe and Asia (Craine et al. 1997, Gylfe et al. 1999). The reservoir of B. burgdorferi ss in the northeastern and upper-midwestern United States includes the white-footed mouse (Peromyscus leucopus) and in some areas eastern chipmunks (Tamias striatus) (Mather et al. 1989, Slajchert et al. 1997, Jones et al. 1998, Piesman 2002). In the far western United States, reservoirs appear to include western gray squirrels (Sciurus griseus) (Lane et al. 2005a) and dusky-footed wood rats (Neotoma fuscipes) (Brown and Lane 1992, Piesman 2002). Borrelia bissettii, another member of the complex in California and Colorado, is maintained in cycles involving wood rats (Neotoma mexicana) and the specialist vector I. spinipalpis (Brown and Lane 1992, Maupin et al. 1994, Piesman 2002, Vredevoe et al. 2004, Brown et al. 2006).
Vertebrate hosts influence transmission cycles in many ways. Most importantly, some hosts serve as a food supply for vector populations and some serve as competent reservoirs of the pathogenic agents. Cervids, including white-tailed deer (Odocoileus virginianus) (Wilson et al. 1990, Fish 1995, Ostfeld et al.
2006), black-tailed deer (Odocoileus hemionus columbianus) (Lane and Burgdorfer 1986; Lane et al. 1994, 2005b), roe deer (Capreolus capreo- lus) (Jaenson and Talleklint 1992, Matuschka et al. 1993), fallow deer (Cervus dama) (Gray et al. 1992, Matuschka et al. 1993), and European red deer (Cervus elaphus) (Gray et al. 1992, Matuschka et al. 1993), are key hosts for the adults of I. ricinus group ticks. Female I. ricinus group ticks each produce as many as 2,500 eggs, and the abundance of these ticks consistently is greater in areas where deer are common (Wilson et al. 1990, Gray et al. 1992, Daniels et al. 1993, Matuschka et al. 1993, Fish 1995, Gern and Humair 2002, Piesman 2006). Deer and other large mammals are important because they sustain large numbers of adult ticks, but ecological maintenance of B. burgdorferi sl depends on the immature stages that feed on competent reservoir hosts.
In addition to those hosts that support ecological maintenance of B. burgdorferi sl, some hosts actually may diminish or disrupt the cycles. The diversity of hosts in a community influences the prevalence of infection in ticks, and therefore the risk of transmission among wildlife and to humans (Ostfeld and Keesing 2000, LoGiudice et al. 2003). Increased diversity of available hosts results in a relatively smaller percentage of the larval ticks feeding on reservoir hosts and fewer nymphs becoming infectious. Thus, the prevalence of infection in both the reservoir hosts and the vectors is expected to be lower when generalist vectors feed on a diversity of hosts that vary in their reservoir competence. Such effects have been dubbed dilution effects because host diversity tends to dilute the prevalence of infection in questing vectors (Ostfeld and Keesing 2000, LoGiudice et al. 2003).
The dilution effect is taken to the extreme when hosts have characteristics that actually kill the pathogens in infected vectors; such hosts are called barrier or zooprophylactic hosts. The prevalence of spirochetes in ticks removed from roe deer, red deer, fallow deer, and wild sheep (Oris ammon) was less than expected based on prevalence in unfed questing ticks (Matuschka et al. 1993). Thus these ungulates actually may have cleared infections in previously infected ticks during feeding, but the mechanism for the “clearing” remained unknown. The prevalence of borrelial infections in I. pacificus from California is generally much lower than I. scapularis in the northeastern United States, and there are several potential reasons for this difference, including differences in the patterns of seasonal activity of immature ticks and slight differences in vector competence (Piesman 2002). However, the key difference may be that more immature I. pacificus feed on zooprophylactic lizards than on reservoir competent hosts (Lane and Loye 1989, Talleklint-Eisen and Eisen 1999, Eisen et al. 2001, Casher et al. 2002, Eisen et al. 2004). Blood of some lizard species contains a heat-labile protein that kills B. burgdorferi (Lane and Quistad 1998); borreliacidal activity of the alternate complement pathway of serum first was demonstrated for western fence lizards (Sceloporus occidentalis) and southern alligator lizards (Elgaria multicarinata) (Kuo et al. 2000). Complement-mediated killing of B. burgdorferi ss and B. bissettii occurs in the serum of both western fence lizards and mule deer, but not in the serum of rodents or birds (Kuo et al. 2000, Ullmann et al. 2003). Mortality of B. burgdorferi ss and B. lonestari also occurred in serum from eastern fence lizards (Sceloporus undulatus) (Grigery et al. 2005). Although lizards are generally not reservoir competent, their impacts on transmission risks are complicated because lizards feed large numbers of immature ticks in some areas (Giery and Ostfeld 2007, Salkeld and Lane 2010, Tijsse-Klasen et al. 2010, Swei et al. 2011). Community dynamics maintaining these cycles can be quite complex, with some host species serving as competent reservoirs, others diluting the prevalence in vectors by diverting their feeding away from reservoir hosts, and still others that have constituents in their blood that actively kill spirochetes inside the fed ticks.
clinical effects Wildlife generally have minimal, mild, or an absence of pathologic disease associated with infections of B. burgdorferi sl. Disease in nonhuman mammals tends to be less severe than in humans; dogs and horses with Lyme borreliosis typically are diagnosed with intermittent fever, swollen lymph nodes, and pain that shifts between leg joints. Disease consistent with some of these signs has been reported in wildlife, but disease in most species of wildlife remains unrecognized or thought to be of minimal to mild severity (Acha and Szy- fres 2001, Brown and Burgess 2001, Schwanz et al. 2011). Although there may be costs associated with energetic losses or behavioral changes of individuals, Lyme borreliosis is not known to cause population level effects in wildlife.
Human cases of Lyme disease manifest in a variety signs and symptoms. Early Lyme disease is often characterized by flu-like symptoms including fatigue, chills and fever, headache, muscle and joint pain, swollen lymph nodes, and a characteristic “bulls-eye” skin rash called erythema migrans (EM). The rash induced by borrelial infection is a flat, non-painful, expanding red ring with a pale center that develops 3 to 30 days after the tick bite (typically 1-2 weeks) (Marques 2010). The EM is distinct from the inflamed, hot, painful, raised rash that develops within minutes or hours after the tick bite from the reaction to tick salivary components around the tick bite; the rapid development of a hot, inflamed, rash does not indicate exposure to spirochetes.
Later stages of Lyme disease progress as the spirochetes become systemic and then sequestered in connective tissues. Signs of rheumatic arthritis are associated with pain, swelling, and erosion of large limb joints, especially the knees; “arthritis” may shift between joints or cause chronic pain in a single joint. In addition, Lyme disease may cause facial nerve paralysis or, more rarely, heart disease. Unfortunately, diagnosis is often difficult because none of these signs is present in all cases; even the characteristic rash is only present in 30-80% of human cases. Late-stage neurologic Lyme disease often affects memory, concentration, and moods. There is also a common late-stage skin condition that occurs in Europe and Asia as a result of infections with B. afzelii (Marques 2010).
special problems No specific special problems are identified.
control No control procedures are established for wildlife. Control of risks of humans acquiring Lyme disease has been achieved by a variety of means. These include personal protection such as wearing appropriate clothing, using tick repellents, routinely checking for ticks, and promptly removing ticks (Piesman 2002, Stafford and Kitron 2002). Other steps include habitat management such as removing brush and leaf litter around homes (Stafford and Kitron 2002), fire (Stafford and Kitron 2002, Padgett et al. 2009), exclusion fences or population reduction of ungulate and other reservoir hosts (Daniels et al. 1993, Stafford 1993, Stafford and Kitron 2002), the use of acaricides to kill ticks in the environment or on hosts (Mather et al. 1987, Daniels et al. 1991, Stafford 1992, Leprince and Lane 1996, Pound et al. 2000, McGraw and McBride 2001, Stafford and Kitron 2002, Hoen et al. 2009), and vaccination of hosts against B. burgdorferi sl or the vectors (Straubinger et al. 2001, Hayes and Schriefer 2002, Tsao et al. 2004, Vrzal et al. 2005, Richer et al. 2011).
Current knowledge gaps regarding control of risks in the northeastern United States were identified recently. These included quantification of risks of exposure at spatial scales relevant to control efforts, patterns of case occurrence in treated and untreated neighborhoods, whether tick population reduction can result in significant risk reduction across a gradient of habitats and characteristics of fragmented forests, the effects of landscape structure on tick-control strategies, knowledge of human use of areas with high acarological risk, and understanding of human acceptance of personal and environmental tickcontrol intervention strategies (Eisen et al. 2012).
Although a Lyme disease vaccine was approved for use in humans in 1998, it was removed from the market in 2002 (Plotkin 2011, Shen et al. 2011). Considerable research has been reported and an effective vaccine (or different vaccines for different regions of the world) might eventually help control risks for people with high exposures to vectors; nonetheless, at the time of writing, that time is yet to come.
Clearly, the complexity of cycles involving different genospecies of B. burgdorferi sl, vector species, and host species make it difficult to recommend simple solutions for control of transmission; rather, we should expect that effective control will remain a moving target and that combinations of control strategies found to be effective in one area may fail to control risks in others.
More on the topic VECTOR-BORNE BACTERIA:
- VECTOR-BORNE VIRAL DISEASES
- NATIONAL VECTOR BORNE DISEASE CONTROL PROGRAMME (NVBDCP)
- Tick-Borne Infections
- BACTERIA
- Segmented Filamentous Bacteria
- RESERVOIRS TYPES USED BY BACTERIA AND VIRUSES
- BACTERIA MAINTAINED BY HOSTS WITH CLINICALLY ACTIVE INFECTIONS
- TICK-BORNE ENCEPHALITIS
- Mosquito-Borne Infections
- TICK-BORNE DISEASES
- Bacteria play a significant role in clinical disease in the rabbit.
- 4 WILDLIFE PATHOGENS WITH ARTHROPOD VECTORS
- INTRODUCTION
- PUBLIC HEALTH CONCERN