CIRCOVIRUS INFECTIONS IN WILD BIRDS
DANIEL TODD
Agri-Food & Biosciences Institute, Stormont, Belfast, UK
Avian circovirus diseases cause feather abnormalities in some avian hosts, most notably psittacine species, but primarily they are causally associated with immunosuppression, which can lead to growth retardation and ill thrift (wasting), and increased susceptibility to secondary pathogens1-2).
Infectious Diseases of Wild Mammals and Birds in Europe, First Edition. Edited by Dolores Gavier-Widen, J. Paul Duff, and Anna Meredith. © 2012 Blackwell Publishing Ltd. Published 2012 by Blackwell Publishing Ltd.
EPIDEMIOLOGY
Information relating to the distribution of circovirus infections of free-living avian species within Europe is confined to single case reports concerning gulls in Sweden(3), starlings in Spain(4) and mute swans in Germany(5). Based on information from racing pigeons and farmed geese and ducks, it is likely that circovirus infections are prevalent within flocks and widespread in avian species, including many wild species.
The recent characterizations of taxonomically distinct circoviruses in different avian species, including pigeons, geese, ducks, gulls, finches, starlings and swans, support the views that novel circoviruses will be recognized in additional avian hosts and that avian circoviruses display high levels of host specificity. Information gathered from studies of psittacine species, pigeons, geese and ducks indicates that most birds become infected as juveniles.
No information is available relating to the influence of climate on the prevalence of avian circovirus diseases. Cir- covirus disease in gulls occurred in summer- autumn in New Zealand(6), whereas deaths (due to salmonellosis associated with circovirus infection? of starlings occurred in winter in Spain(4).
There is no information as to whether circoviruses use insect vectors, but this is considered unlikely.Cross-infection between closely related avian species remains a possibility; however, given the high degree of host-specificity, it is unlikely to occur between unrelated species. Cross-infections between free-living birds such as ducks or geese and their farmed counterparts are considered possible.
Investigations with psittacines and racing pigeons indicate that circoviruses can replicate in the intestinal tract and can be excreted for weeks to months after infection. Because circoviruses are highly resistant to inactivation, environmental contamination is high and transmission by ingestion of infected material is likely. The detection of circoviruses in the reproductive tissues (PiCV) of adult birds and in embryos (PiCV, BFDV) suggests that vertical transmission of avian circoviruses is possible.
PATHOGENESIS, PATHOLOGY AND IMMUNITY
Information relating to pathogenesis is mainly confined to psittacines, racing pigeons and farmed geese. It is probable that most birds become infected within the first few weeks with horizontally acquired virus, although evidence from pigeons indicates that infections may also be acquired vertically. There is no information on the infectious dose required, mainly because of the absence of cell culture propagation systems for estimating virus infectivity.
Ingestion of virus-contaminated material is likely to lead to infection of the intestinal tract, a tissue in which the circoviruses of psittacines, pigeons and geese have been detected. Circovirus viraemias have been demonstrated in psittacines, pigeons and geese, and this is the probable mechanism by which circoviruses spread to other organs and tissues. In clinically affected birds, circoviruses can be detected in a wide range of tissues. For example, PiCV has not only been detected in lymphoid tissues including the bursa of Fabricius and spleen, but also in a wide range of other tissues, including the intestine, respiratory organs (trachea, pharynx, lung), kidney, liver and reproductive tissues (ovary and testes).
Owing to their very limited protein-coding capacities, circoviruses are heavily dependent on the host cell’s replication machinery and probably replicate best in rapidly dividing tissues such as epithelial cells. In diseased birds, circovirus inclusions are commonly found in the cytoplasm of macrophages (Figure 4.1). Such inclusions probably result from phagocytosing activity.Most avian circovirus infections are likely to be subclini- cal, probably involving low levels of virus replication in a limited tissue range. Clinical manifestations including ill
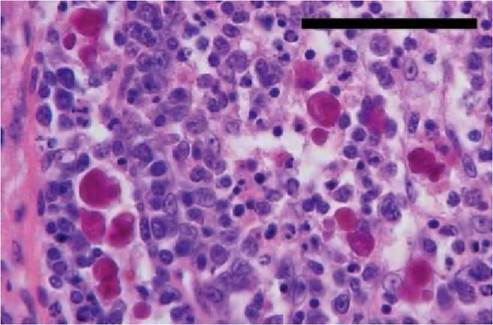
FIGURE 4.1 Bursa of Fabricius from a juvenile herring gull showing depletion of lymphocytes, eosinophilic inclusions in both cortex and medulla of the bursal follicle. Haematoxylin and eosin. Scale bar = 50 μm. (See Smyth et al., 2006)(3).
thrift, and feather abnormalities usually arise when high levels of virus are detectable within tissues. High virus levels may correlate with lymphocyte depletion and with the resulting immunosuppression. Studies with pigeons and geese suggest high levels of virus in tissues may take several weeks to develop. In chronically infected psittacine species, there is progressive feather loss and occasional beak deformity. Although cases of clinical remission have been observed, fatal outcomes, often resulting from secondary infections exacerbated by the circovirus-induced immunosuppression, are frequent. In non -psittacines the circovirus- induced immunosuppression causes less-specific clinical signs, including ill thrift and weight loss, with the clinical manifestations observed again being attributable to secondary opportunistic infections. For example, in the case of a southern black- backed gull (Larus dominicanus) the pathological findings were consistent with the possibility that a circovirus-induced immunosuppression allowed an overwhelming secondary infection with Aspergillus spp.(6).
A number of factors have been proposed to explain why some infections result in disease, whereas others remain subclinical. These include the route and age at which the birds are infected, the levels of maternal antibody, virus strain, infectious dose, and presence of co-infecting pathogens. Antibody responses to circovirus infections have been detected in a limited range of avian species, including psittacines, geese and pigeons. Experiments with psit- tacines have shown that maternally derived antibody is protective against a disease challenge.Feather loss and beak deformities are the principal lesions associated with psittacine beak and feather disease, the most studied avian circovirus disease. Feather dystrophy and loss are usually symmetrical, with changes in the feathers including retention of sheaths, haemorrhages within the pulp, cavity fracture of the proximal rachis and failure to exsheath also being observed. Lymphoid depletion including bursal atrophy can be a feature of avian circovirus diseases, but these lesions can also be due to infections with other viruses. Many of the gross lesions observed in non-psittacines are attributable to secondary pathogens. At the microscopic level, affected birds show lymphocyte depletion and infiltration of monocytes. Basophilic globular or botryoid inclusions can also be observed in the cytoplasm of macrophages found within lymphoid tissues such as the bursa of Fabricius (Figure 4.1). Thin-section electron microscopy has shown that these inclusions comprise paracrystalline arrays of tightly packed circovirus particles (Figure 4.2).
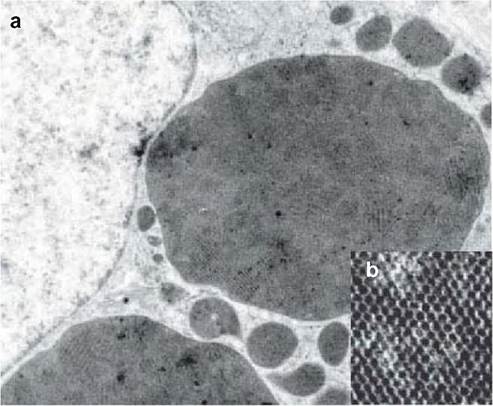
FIGURE 4.2 Thin-section electron microscopy performed with bursa of Fabricius tissue from a circovirus- infected psittacine species. Electron- dense inclusions containing paracrystalline arrays of tightly packed virus particles were observed in the cytoplasm of macrophage cells and are shown at low (a) and high (b) magnification.
Image provided by Dr Joan Smyth, AFBI- Stormont, Belfast (See Smyth et al., 2006)(3).CLINICAL SIGNS AND TREATMENT
Feather abnormalities are a common manifestation of BFDV infections in psittacines, and have also been observed in non-psittacine species, including Senegal doves (Streptopelia senegalensis) infected with a PiCV variant, pigeons, Australian ravens (Corvus coronoides')i'τ,, finches, ducks and geese. Non-psittacine species frequently show non-specific clinical signs, including weight loss and ill thrift, with specific signs being attributable to secondary pathogens.
It may be possible to rehabilitate individual birds that are found sick by treating secondary infections. Clinical remissions have been observed with some psittacines and with Australian ravens, and infected psittacines have been successfully treated with avian gamma interferon(8).
DIAGNOSIS
Infections with circovirus-like viruses were originally diagnosed in pigeons, gulls and farmed geese based on the detection of characteristic, basophilic globular or botryoid inclusions, which were first described in psittacines infected with BFDV. Electron microscopic techniques have also been used to detect paracrystalline arrays of small (14— 17nm) tightly packed, virus particles (Figure 4.2) within the inclusions and circovirus-like particles in tissue homogenates prepared from pigeons, geese, ducks and pheasants. However, owing to the absence of surface structure and variation in the accuracy with which small virus particles can be measured, definitive diagnoses should not be based on negative contrast electron microscopy alone.
So far, growth of most avian circoviruses in cell culture has not been described. This has meant that virus culture cannot be relied on for diagnosis, and it has also greatly restricted the ability to produce virus-specific antisera for use in antigen-detecting diagnostic procedures such as immunohistochemistry and cryosection/ impression smear fluorescent antibody tests.
Molecular approaches are used for virus characterization and diagnosis. Degenerate primer-based polymerase chain reaction (PCR) methods and other novel DNA-amplifying methods have been used to amplify sub-genomic and full-length genome fragments specific to the majority of circoviruses recognized to date. Phylogenetic analyses based on sequence comparisons have then been used to establish whether the circovirus detected represents a new genus member. Knowledge of the circovirus sequences and the availability of cloned genome fragments have facilitated the development of diagnostic methods based on DNA detection, including in situ hybridization (ISH), dot blot hybridization (DBH) and PCR. PCR tests have been described for the specific detection of BFDV, PiCV, GoCV, DuCV, StCV and SwCV. Applications of these tests have shown that infections are prevalent and likely to be widespread. However, PCR detects subclinically infected birds as well as birds that are clinically affected by circovirus. Therefore, owing to its very high sensitivity, PCR may be of questionable use in disease diagnosis. Although ISH is less sensitive, its ability to demonstrate substantial amounts of virus DNA within lymphoid tissues may prove to be more valuable for diagnosing avian circovirus diseases (Figure 4.3). With birds infected with PiCV, GoCV and GuCV, virus DNA was detected by ISH in a range of tissues, many of which — for example liver — were not showing histological changes. A digoxigenin-labelled ISH probe prepared from a cloned PiCV genome fragment containing part of the rep gene has been shown to cross- r eact with GoCV, CaCV and GuCV. As such, this may prove to be a useful reagent for the detection of novel circoviruses in other avian species(3).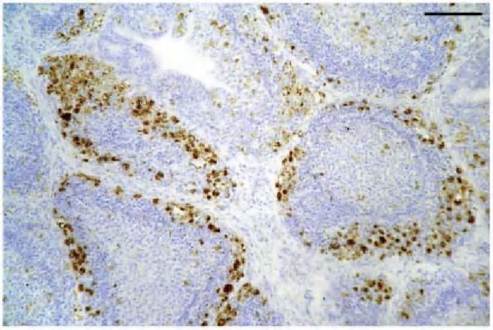
FIGURE 4.3 ISH showing positive labelling of circovirus DNA (stained brown) in the bursa of Fabricius from a juvenile herring gull. Infected cells are most numerous in the cortex of the follicles. ISH counterstained with haematoxylin and eosin. Scale bar = 100 μm. (See Smyth et al., 2006)(3).
PCR is likely to prove to be the most suitable test for screening populations for circovirus infections. Investigations with pigeons have shown that cloacal swabs taken many months post infection, are PCR- positive, and this may be the case for other avian species. Therefore, cloacal swab samples collected from captured juvenile wild birds or faecal samples collected from nesting sites will probably contain PCR- detectable virus resulting from subclinical and clinical infections. Serological testing is likely to be less useful, because antibody- detecting tests are available for very few avian circoviruses.
MANAGEMENT AND CONTROL
To date in Europe, outbreaks of circovirus diseases in free- living birds have been limited to those in occurring in gulls, starlings (Sturnus vulgaris) and swans, in which cir- covirus was detected in birds that were found dead. Management and control of circovirus disease in wild bird populations are impractical on the basis that it is difficult to administer vaccines or therapeutics to free-living birds.
PUBLIC HEALTH CONCERN
Avian circoviruses are highly host-specific, and infections of humans are considered highly improbable.
SIGNIFICANCE AND IMPLICATIONS FOR ANIMAL HEALTH
Although it is known that circovirus infections of farmed geese and ducks are common and widespread, at present there is no information about: i) the prevalence of GoCV and DuCV infections in free-living birds; and ii) whether the circoviruses that infect the free-living birds are identical to those that infect the farmed birds. On this basis, the possibility of cross-infections occurring between the farmed and free-living birds cannot be discounted. Crossinfections with BFDVs, which are known to infect a range of psittacine species, will occur between captive and free- living birds unless sufficient biosecurity is maintained.
More on the topic CIRCOVIRUS INFECTIONS IN WILD BIRDS:
- CIRCOVIRUS INFECTIONS IN WILD BIRDS
- CIRCOVIRUS INFECTION IN WILD BOAR
- 7 DISEASES BY CLINICAL PRESENTATIONS, BIRDS
- 5 Appendices
- PORCINE PARVOVIRUS INFECTION IN WILD BOAR
- HERPESVIRUS INFECTIONS IN WILD MAMMALS
- Macrorhabdus Ornithogaster infection in BIRDS
- RELAPSING FEVER SPIROCHAETE- Associated disease in birds and MAMMALS
- Brachyspira infections in birds
- MYCOPLASMAS OF BIRDS