Disorders of Skin
Mechanical Muzzle Alopecia
Hair loss in the muzzle region occurs occasionally in laboratory mice due to mechanical denuding from improperly constructed openings for feeders or watering devices.
This must be differentiated from barbering.Alopecia Areata in C3H Mice
Aging C3H mice develop irregular, diffuse alopecia of the ventral and dorsal trunk, which closely mimics human alopecia areata. Hair loss increases with age, particularly after 6 months of age, in both males and females. Microscopic examination of affected skin reveals densely packed anagen follicles with dystrophic hair formation, “melanin incontinence,” interfollicular epidermal thickening, and perifollicular mononuclear leukocyte infiltrates.
Alopecia of B6 Mice
This extremely common multifactorial syndrome in B6 mice is fundamentally a behavioral disorder. It is influenced by husbandry factors, such as premature weaning and diet. The alopecia is associated with barbering or trichotillomania but becomes secondarily inflammatory in character. B6 mice are prone to Myobia hypersensitivity dermatitis, which can significantly contribute to the syndrome. Immune complex vasculitis and primary follicular dystrophy have been proposed as underlying factors. These factors collectively predispose B6 mice to necrotizing dermatitis, which is driven by opportunistic infection with Staphylococcus and Streptococcus. Ulcerative dermatitis is a major life-limiting disease, with mice succumbing prematurely to systemic amyloidosis and atrial thrombosis. See sections on Staphylococcus, Streptococcus, ectoparasitism, behavioral disorders, amyloidosis, and atrial thrombosis.
Clown Mouse Syndrome
This rare syndrome is seen among weaning-age mice, in which one or the entire litter may be affected. Typically, mice are runted and have general alopecia, with varying amounts of normal-appearing hair on the head, neck, and thorax (Fig.
1.108). There is also often hyperkeratosis of the affected hairless skin. Affected mice have ongoing severe systemic disease or have recovered, resulting in temporary cessation of normal hair growth cycling. Hair growth cycles extend from head to tail, so that regrowth appears first around the head and progresses posteriorly. Thus, the amount of hair varies and may extend over the neck and thorax in some animals. Clown mouse syndrome has especially been associated with both natural and experimental mouse hepatitis virus infection.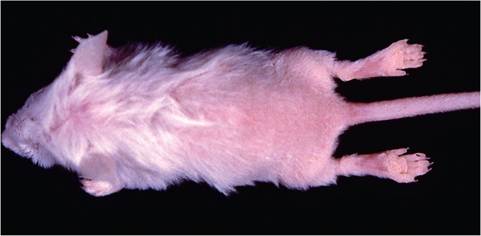
FIG. 1.108. Clown mouse syndrome in a young mouse due to arrested hair growth following mouse hepatitis virus infection.
Ear Gangrene and Notching
Erosive inflammation and necrosis of the edges of the ear pinnae, with formation of notches in healed lesions, has been described in Swiss mice, but the authors have observed the lesion frequently in C3H mice as well. The etiology of this disorder is not known, but it is assumed to be environmental. Lesions begin as erythema, progress to erosions with serous exudation, and eventually develop into necrosis of the ear. Histopathology reveals hyperkeratosis and underlying chronic inflammation. Severely affected mice may develop ulcerative dermatitis of the neck and shoulders.
Auricular Chondritis
Both mice and rats are prone to development of bilateral thickening of the ear pinnae (Fig. 1.109) due to underlying chronic granulomatous inflammation and degeneration of cartilage. Lesions may be diffuse or nodular in appearance. Lesions are associated with metal identification tags, which are believed to stimulate an autoimmune chondritis. The autoimmune basis is underscored by the development of similar lesions in the contralateral, untagged ear, and induction of the lesion by immunization with type II collagen. Lesions may stimulate formation of squamous cell carcinomas.
Ringtail
Low ambient humidity is associated with “ringtail,” or annular constrictions of the tail and occasionally digits in infant mice, resulting in edema of the distal extremity and dry gangrene.
Hairless strains of mice are prone to skin problems as adults, which may be manifest as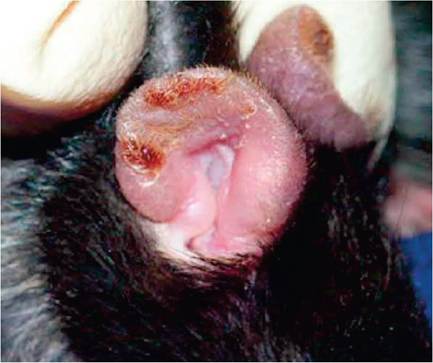
FIG. 1.109. Auricular chondrosis in a B6 mouse. This inflammatory syndrome is believed to have an autoimmune basis, can be experimentally induced by collagen immunization, and often arises naturally in response to ear tags. (Source: Buckner, J.H., David, C.S. & Bradley, D.S. (2006) Mice expressing HLA-DQ6alpha8beta transgene develop polychondritis spontaneously. Arthritis Research and Therapy 8:R134. Reproduced under the Creative Commons License http:// creativecommons.org/licenses/by/4.0/)
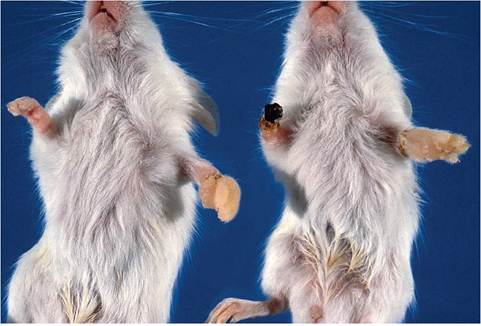
FIG. 1.110. Edema and gangrene of digits and feet of suckling mice associated with inadvertent winding of cotton fibers around digits and limbs. (Source: Percy et al. 1994. Reproduced with permission from American Association for Laboratory Animal Science.)
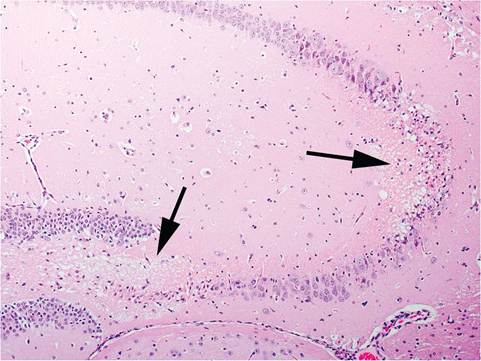
FIG. 1.111. Post-ictal neuronal necrosis (arrows) in the hippocampus of an FVB mouse.
inflammation and gangrene without the classic ringtail antecedent stage. The nature of the predisposing factors and the precise pathogenesis of ringtail have yet to be resolved but are thought to be related to low (less than 40%) relative humidity and high (greater than 80°F) temperature. Lesions must be differentiated from behavior-related dermatitis of the tail.
Sloughing of Extremities Due to Cotton Nesting Material
Necrosis and sloughing of digits and feet in suckling mice have been associated with infarction due to the wrapping of absorbent cotton (cotton wool) nesting material around one or more legs or digits (Fig. 1.110).
Frostbite
Rarely, mice may develop gangrene of their limbs due to frostbite. Nude mice are especially prone to this syndrome, which is associated with cold exposure of shipping boxes during air transportation.
Mammary Hyperplasia
Virgin female FVB/N mice develop mammary gland lobuloalveolar hyperplasia, with secretory product in alveoli and ducts. These changes increase with age and are correlated with the presence of proliferative lesions of prolactin-secreting cells in the pars distalis of the pituitary. In addition, FVB/N mice are prone to a high incidence of pituitary adenomas and mammary tumors in aged multiparous females. Persistent mammary hyperplasia, without involution, is especially prominent in multiparous FVB/N females and may be present in the absence of pituitary changes.
Disorders of the Central Nervous System and Sensory Organs
Seizures
Several strains of mice, particularly DBA/2 mice, but also SJL, LP, and FVB mice, are prone to audiogenic
(high-frequency) seizures. Susceptibility in DBA mice wanes with age. B6, C3H, and BALB/c mice tend to be resistant, but some BALB mice may be susceptible at old (>400 days) age. Seizures progress after a short latency period following sound stimulation. Mice manifest an explosive bout of frenzied running, followed by clonic seizures with violent kicking movements. A tonic phase ensues with rigid extension of limbs and flattening of ear pinnae. During this phase, respiratory arrest may occur. Individual mice may not progress through all phases. Other forms of sensory stimulation, including alteration of equilibrium, can induce seizures in some mice, such as EL mice. Some populations of FVB mice are highly prone to audiogenic seizures, which have been elicited by tattooing, hair clipping, and fire alarms. Seizures often occur at night, resulting in unexplained deaths. Seizures are characterized by facial grimace, chewing automatism, whole-body tics, ptyalism, and clonic convulsions. At necropsy, there is neuronal necrosis in the cerebral cortex, hippocampus, and thalamus, especially in the organized pyramidal cell layers of the hippocampus (Fig. 1.111). Mice that have endured multiple convulsive events may have generalized gliosis.
Acute coagulation necrosis of centrilobular hepatocytes may also be present in some mice. Other common observations at necropsy include enlarged and flaccid urinary bladders, distended gall bladders, enlarged adrenals, and increased brain weights. A general consequence of seizures in male mice may be ejaculation, retention of urethral plugs, and obstructive uropathy.Hypocallosity
Forebrain commissure defects, with aplasia of the corpus callosum, are common (up to 70%) in BALB/c, 129 (including 129/J, 129/Sv, 1229/ReJ, and 129/Ola), and some other less common strains of mice. This feature makes the affected animals poor candidates for learning behavior research, and hypocallosity has been mistaken as a “phenotype” in GEMs derived from 129 ES cells.
Hydrocephalus
Internal hydrocephalus is relatively common among C57BL mouse strains. It becomes clinically significant at weaning, when mice are unable to independently eat and drink. Mice present with domed skulls and are usually runted and dehydrated. Recently, it has been found that high levels of Cre recombinase expression in neuronal precursor cells can lead to reduced neuronal proliferation and apoptosis, with defects in ependymal lining, lamination of the cortex, microencephaly, and communicating hydrocephalus. This phenomenon was noted in 3 different nestin-Cre transgenic lines of mice.
Central Nervous System Miscellany
Vacuolation of the white matter is a common fixation artifact in mouse brains. Artifactual vacuoles can be differentiated from antemortem vacuolation by their distinct outlines, lack of cellular debris, and lack of gliosis. Multilaminated mineralized concretions are common in the thalamus of old mice. Developmental midline malformations are relatively common, including hamartomas, choristomas, teratomas, epithelial inclusion cysts, and adipose tissue (lipomas). A number of other artifacts and lesions have been described.
Cochlear Degeneration
Age-related hearing loss is common in many strains of inbred mice.
Its basis is genetically complex, with different mutant genes, modifier genes, and pathogenesis, depending upon mouse strain. At least 8 mapped loci and a mitochondrial variant are known to contribute to age-related hearing loss in mice. A survey of 80 inbred mouse strains has revealed 18 strains that have significantly elevated auditory-evoked brain stem response thresholds by 3 months of age, including 129P1/ReJ, A/ J, DBA/2, and NOD/Lt mice, and others that have elevated thresholds at older ages, including B6 and BALB/c mice. High-frequency losses occur first, with cochlear hair cell loss progressing from the base to the apex. Outer hair cells are affected before inner hair cells, and ganglion cells degenerate as a consequence of hair cell loss.Vestibular Syndrome
A significant manifestation of polyarteritis (see “Polyarteritis”) is vestibular syndrome. Head tilt, circling, and more severe manifestations of vestibular disease are common clinical signs. This syndrome occurs in a number of mouse strains in the absence of detectable viral and bacterial causes of otitis. The internal and middle ear structures are normal, but careful examination of surrounding tissues will reveal active necrotizing and/or inflammatory changes in medium-sized arteries (Fig. 1.112). It is typically found with concomitant involvement of arteries in other tissue, particularly
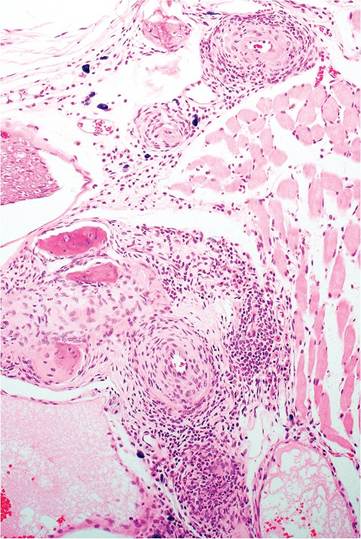
FIG. 1.112. Chronic arteritis near the middle and inner ear. Arteritis occurs in multiple organs in adult mice with polyarteritis, but involvement of vessels in the head may result in vestibular signs.
coronary arteries. Other causes of vestibular syndrome include otitis, neoplasia, and spontaneous unilateral brainstem infarction, which has been reported to occur in Swiss mice.
Corneal Opacity
Corneal opacities have been observed in a variety of strains of mice. Opacities are characterized by acute to chronic inflammatory changes of the corneal epithelium and anterior corneal stroma, including acute keratitis with corneal erosion to ulceration, vascularization of the corneal stroma, and mineralization of corneal basement membranes. In some cases, the problem can be alleviated by more frequent cage cleaning, and it has been concluded that an environmental factor, such as ammonia, may play a role in the development of the disease.
Blepharitis and Conjunctivitis
Several common inbred strains of mice are prone to develop blepharitis and blepharoconjunctivitis, including 129P3/J, A/HeJ, BALB/cJ, BALB/cByJ, and CBA/J mice. Suppurative conjunctivitis with ulcerations at the mucocutaneous junction has been observed in 129, BALB, and other strains of mice. Abscessation of the meibomian glands may also occur. A variety of bacteria have been isolated from affected conjunctivae, including Coryne- bacterium, coagulase-negative Staphylococcus, and P. pneumotropica. These are likely to be opportunistic infections, and the specific etiopathogenesis is unknown. C57BL mice are also prone to develop blepharoconjunctivitis in association with microphthalmia, and mutant mice that have premature opening of eyelids are prone to eye trauma and blepharoconjunctivitis.
Microphthalmia and Anophthalmia
This syndrome is relatively common and unique to various types of C57BL mouse, with females more prone to lesions than males. Mice usually have asymmetric microphthalmia or anophthalmia, most often involving the right eye.
Retinal Degeneration
Homozygosity of the Pde6brd1 allele is common among both inbred and outbred mice, resulting in retinal degeneration and blindness at an early age. Retinal degeneration is a strain characteristic of several common inbred strains of mice, including C3H/He, CBA, FVB, SJL, SWR, and others. It also occurs in most outbred Swiss and nonSwiss albino stocks and may be observed in some populations of wild mice. A study examined the prevalence of this lesion among outbred albino mice and found retinal degeneration to be very common, but there was marked variation in prevalence, depending upon the mouse stock. Notably, retinal degeneration was not found in Crl:CD-1(ICR)BR, HsdWin:CFW1, and Hsd:NSA(CF-1) mice. NIH Swiss and Black Swiss mice are also affected. Mice are born with normal-appearing retinas and develop normal photoreceptors that rapidly undergo apoptosis, beginning in the third wk of life. Affected mice become totally blind. Microscopic changes include absence or degeneration of the rods, outer nuclear layer, and outer plexiform layer (Fig. 1.113). Active
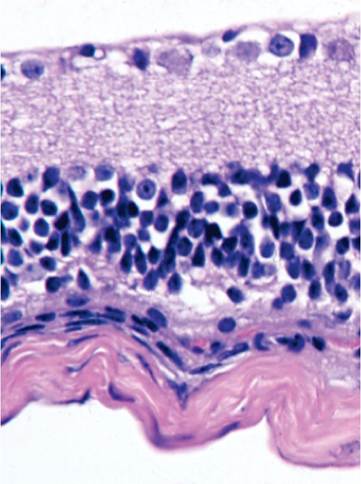
FIG. 1.113. Retinal degeneration in a C3H mouse that is homozygous for the rd1 (rodless retina) allele. There is loss of the outer plexiform and bipolar cell layers.
degenerative changes can be encountered in young mice, but the lesion evolves rapidly within a few weeks of weaning. Unaware investigators have often been duped by blind mice that fail to perform normally in behavioral studies.
More on the topic Disorders of Skin:
- Disorders of Skin
- Agrawal M.. Textbook of Pediatrics. 3rd ed. — CBS Publishers,2025. — 973 p., 2025
- Inspection
- Physical Examination
- PORPHYRIA
- Peripheral Vascular Disease
- Genitourinary Infections
- THE CLOSTRIDIAL DISEASES
- CLINICAL EVALUATION OF GIT DISEASE
- HYPERTHERMIA