Postmortal Examination (PME)
Of all the available postmortal diagnostic techniques, recognizing the lesions caused by an M. bovis infection in the various species remains one of the most important, and when correctly done, one of the most Se, Sp, and cost-effective tests with which to diagnose BTB (Rogers et al.
1980; Whipple et al. 1996; Liebana et al. 2008; Nunez-Garcia et al. 2018). The ability to recognize and detect the lesions determines the success and quality of meat inspection in abattoirs and when examining incidental deaths as part of passive surveillance for BTB.Both in countries where abattoir surveillance is used to monitor the absence of the disease, and in resource-poor countries where it is the only affordable method for BTB detection, it is thus critical for investigators to be fully acquainted with the physical appearance and distribution of the lesions and the inter- and intraspecific variation that may occur both in the macroscopical and histopathological appearance of the lesions and in their distribution. These may differ substantially, and unless examiners are aware of these characteristics, they may easily miss lesions in infected carcasses. If they do not know where to look, and what to look for, they will in all likelihood miss the lesions and the diagnosis. The least that can be expected of veterinarians and meat inspectors involved in this type of investigation is that they know where the relevant regional lymph nodes are located, and the variation in their normal appearance. They should also have an understanding of the inter- and intraspecific appearance and the variation in the lesions caused by M. bovis and other relevant mycobacteria that may infect cattle, small-stock and camels, and free- ranging African wildlife species.
The pathology of BTB is well documented in a number of older textbooks (Francis 1958; Jubb et al. 1993), and it appears that in those countries from which the disease has been eradicated, the lesions assume a different characteristic macroscopical appearance as they are mostly early lesions.
In those countries, the advanced disease, with severe generalized lesions, and the full scope of the pathology of BTB is rarely seen. This should be kept in mind in African countries in which the disease has never been controlled, as the lesions and general manifestation of BTB may be substantially different (Domingo et al. 2014). This may have an influence on the ability to recognize the lesions and to detect diseased carcasses. It is important to monitor the appearance of lesions in different countries, as it will also give an indication of the nature of the disease and may influence control strategies. Further issues such as latent infections, the presence of no visible lesions (NVLs) in tested herds, and the distribution of lesions that can be used to improve the ease and accuracy of their detection should also be considered (Cassidy 2008).Postmortal examination for diagnosing BTB is a powerful tool, if applied correctly. However, investigators must be adequately trained to recognize the lesions, and do their work with the necessary diligence and meticulous attention to detail, if the results are going to be reliable and of any value to the regulatory authorities, and the public at large. Regular monitoring and quality control of the processes should be part of the activities included in control programs.
9.6.1 The Pathology of BTB
Cattle Bovine TB manifests as a chronic, focal or multifocal, granulomatous caseous-necrotic inflammatory process that primarily localizes in the lungs, and their draining lymph nodes, but depending on the portal of entry, may occur in a number of other organs and lymph nodes (Domingo et al. 2014). The distribution of lesions differs according to various reports, and these differences may be a reflection of the different routes of infection by which cattle contract the disease. Generally, it was believed that most infections in cattle were contracted by inhalation, but it now appears that per os infection with localization in the palatine tonsils (in about 20% of cases) may play a significant role in the way in which BTB is contracted.
Exudate, including the mycobacteria present in the lesions in the tonsils, drains into and causes lesions in the medial retropharyngeal lymph nodes, from where the bacteria disseminate to cause lesions in the lungs and elsewhere (Domingo et al. 2014).Following infection, lesions develop in the organ in which it localizes and then extend to the regional lymph node. When granulomas in both these sites are visible, it is referred to as a complete primary complex; lesions in only one of the sites, usually the regional lymph node, are known as an incomplete primary complex. In one study, single lesion occurred in 66% of infected carcasses (Corner et al. 1990), but in others on average, 1.7 lymph nodes were affected in a group of 61 tuberculous animals (Liebana et al. 2008). Percentages of the occurrence and distribution of lesions differ. In carcasses examined by Corner et al. (1990), 29.4% of the lesions were found in the medial retropharyngeal nodes, 28.2% in the mediastinal nodes, 18.0% in the tracheobronchial nodes, 8.0% in the lungs, 2.9% in the mesenteric nodes, 2.4% in the parotid node, and 2.4% in the caudal cervical node. These sites should be the primary focus of inspection to detect infected animals with visible lesions. In some of the animals in which lesions occurred in the abdominal nodes, they were the only ones affected. Lesions are often found in the palatine tonsils (Neill et al. 2001), and they are common sites from which positive cultures can be obtained. They are often the location of non-visible lesions. The issue of no visible lesions (NVLs), i.e., those TST-positive animals in which no lesions can be detected, is an important one. As many as 10% of TST-positive reactors may contain no lesions, and 30% of TST-negative, in-contact animals in one study of a BTB-positive herd, contained NVLs detected by culture following postmortal examination (Gavier-Widen et al. 2009). It is critical to keep in mind that not being able to find lesions at necropsy does not mean that the animal is not infected with M.
bovis.Infection by inhalation normally causes the development of lesions in the lower respiratory tract where they tend to localize in the dorso-caudal portion of the caudal lung lobes, although, to a lesser extent, lesions may be present in all the other lobes of the lung (Liebana et al. 2008). In general, lesions are more commonly found in the thoracic cavity, then in the head, and lastly in the abdomen. When examined with care, pulmonary lesions may be found in up to 70% of animals, and 1.5 lobes on average contain lesions. It is normally easier to find small lesions in the regional lymph nodes compared to the lungs.
Tuberculous lesions on the serosal surfaces manifest typically as multiple, single to confluent clusters of sessile or pedunculated nodules resembling pearls and become markedly calcified (Figs. 9.6 and 9.7). Serosal lesions may proceed to develop a diffuse, caseous serositis with the development of large caseous plaques adherent to the thickened serosal surfaces (Neill et al. 2001).
Conceivably, single lesions could occur anywhere in the body, a situation that makes detection of all lesions a tedious process, and these abnormally located lesions are usually not detected during the course of routine meat inspection. Single lesions also occur unexpectedly in lymph nodes such as the sub-iliac, which is not one of the target nodes for routine meat inspection.
The lesion caused by an M. bovis infection is typically a focal granuloma, manifesting as irregular firm, white or yellowish nodules varying from 2 to 20 mm or more in diameter. They are characterized by the presence of an accumulation of
Fig. 9.6 Tuberculous peritonitis. Note the typical pearl-like granulomas that are often markedly calcified
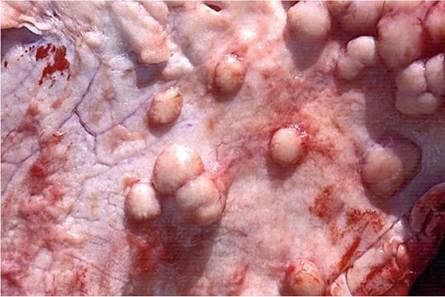
Fig. 9.7 Sessile tuberculous granulomatous plaques on the pleural surface that can easily be confused with mesotheliomas
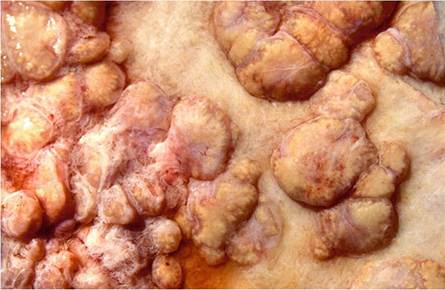
centrally located caseous necrotic material that may or may not become inspissated and calcified, and may undergo liquefaction usually in association with the presence of neutrophils.
Liquefaction of the exudate creates a lesion that is easily confused with an abscess with a fluid purulent content, caused by a variety of pyogenic organisms. A rim of an active granulomatous response and an outer capsule surround the granuloma. Lesions can persist with or without progression, or they can heal and disappear. In cattle experimentally infected with M. bovis, the lesions are classified into four categories according to their stage of development (Cassidy et al. 1998; Wangoo et al. 2005). This then, although lesions following experimental infection may differ from those in natural cases, is the usual concept of the appearance of lesions expected in animals suffering from BTB. The lesions, however, may differ substantially in size and appearance in naturally infected cattle due to attributes that may be dependent on the breed of cattle, the age and gender of the animal, virulence of the mycobacterial strain, the immune status of the animal, and the stage of the disease (Domingo et al. 2014). In the advanced disease, lesions may vary substantially and are inconsistent in appearance with those in experimental animals or in the early stages of the disease.A common characteristic of the lesions of BTB in the various species, including cattle, is the substantial variation that occurs between animals in their macro- and histopathological appearance (Neill et al. 2001; Ameni et al. 2007). In cattle, the lesions are characteristically yellowish-white nodules containing a dry gritty content, and may occupy the entire tissue space contained by the lymph node’s capsule.
Lesions in the lymph nodes vary in appearance and may consist of ill-defined foci of necrosis with a predominantly neutrophil infiltrate, to clearly encapsulate granulomas with calcified caseous necrotic centers. Individual lesions are rarely severely fibrosed, and they lack the thick fibrous capsule that characterizes the so-called “typical tubercle.” Occasionally lymph nodes with the appearance of large encapsulated abscesses containing a thick yellowish to greenish purulent exudate may be seen (Whipple et al.
1996).Single lesions are commonly seen in the lung (Fig. 9.8), but they can be multiple and confluent. In generalized cases, multiple embolic granulomas may be scattered throughout the lungs. In many instances, a single, small granuloma is the only lesion seen, but in individual cases, multiple granulomas of up to 70 mm in diameter and scattered throughout the lungs may be present. In advanced cases, the lesions may obliterate large portions of the lungs, more often in the caudal lobes, and develop into a lobar pneumonia, but with retention of the interlobular septa (Buddle et al. 1994).
Local lesions may expand substantially over time and may destroy the normal structure of the organ. Thus, the expanding inflammatory process may destroy all the normal tissue of a lymph node and cause substantial irregular enlargement of individual nodes. In the lung, the lesions expand to affect whole lobules that may by extension, draining of the exudate via the airways, or dissemination of mycobacteria via the blood or the lymphatic vessels, spread to adjacent or distant lobules. In this way, large portions of the lungs may become affected. Multiple lesions may become confluent, and in advanced cases, the necrotic portions may undergo liquefaction and
Fig. 9.8 A small cluster of focal, pyogranulomatous lesions in the lung resembling abscesses
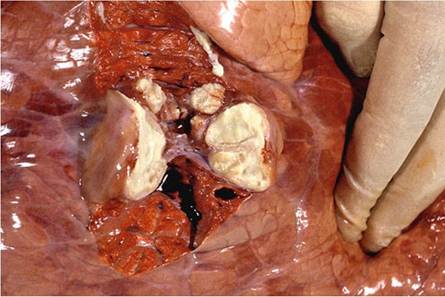
cavitation. These cavities remain small and are limited to lobules because of the very rigid interlobular septa in cattle that persist and limit further expansion of the lesions (Domingo et al. 2014).
Depending on the immune status of an infected animal, dissemination of the infection may occur, either early during the course of the infection or at a later stage, a process that is, respectively, referred to as early or late, postprimary dissemination (or embolic spread) via the blood vascular or lymphatic systems. This type of dissemination usually results in the development of miliary granulomatous lesions in many organs and tissues throughout the body. The lesions in the lungs and on serosal surfaces of the thorax, the abdomen, and the pericardium are more commonly seen. Although they may initially be small, miliary nodules may become confluent and in this way develop into sizeable lesions. In generalized cases, lesions may also occur in the liver, kidneys, udder, and meninges (Neill et al. 2001).
For diagnostic purposes, lesions that macroscopically resemble those caused by mycobacterial infections should be confirmed histopathologically and/or by culture, as only a presumptive diagnosis can be made following the detection of the so-called “typical macroscopic lesions” (Liebana et al. 2008). There are numerous causes of disease that produce lesions with a similar macroscopical appearance to those caused by the mycobacteria. These include organisms of the NocardiaZStreptomyces spp. group, those causing actinomycosis and actinobacillosis, and Staphylococcus, Dermatophilus, Corynebacterium, and Pseudomonas spp. Additionally, hydatid cysts, squamous cell carcinoma, and fungal and foreign body granulomas produce lesions with a similar appearance. In certain instances, what appears to be just an abscess, can turn out to be a lesion caused by M. bovis of which the contents are liquefied. Confirming the diagnosis at least by histopathology, but preferably by culture, is of particular importance when dealing with single animals originating from herds with an unknown BTB history. Not making a diagnosis of BTB under those circumstances could have disastrous consequences.
Histopathologically, lesions develop from an initial accumulation of macrophages, epithelioid cells, neutrophils, and a few Langhans-type giant cells. In time, the central portion undergoes caseous necrosis following which the exudate becomes inspissated and develops varying degrees of mineralization. A typical active granulomatous inflammatory reaction, a peripheral cuff of lymphocytes, and finally a capsule that may not be fully developed, surround the lesion. Varying numbers of acid-fast mycobacteria may be detected in the cellular debris, and in the cytoplasm of the Langhans giant cells and epithelioid cells. Many of the lesions are paucibacillary and histologically contain few or no detectable mycobacteria. It must be again stressed that the lack of detectable acid-fast mycobacteria in histological sections is not a reason to rule out a diagnosis of tuberculosis (Domingo et al. 2014). In addition, only a presumptive diagnosis of BTB can be made on the basis of detecting acid-fast bacteria (AFB) histologically since it is not possible, based on their morphology, to identify either the species or the genus of the acid-fast organisms in histopathological sections (Corner 1994).
9.6.2 Small Stock and Camels
Camels Limited data are available about the disease in dromedary camels (Camelus dromedarius), and tuberculosis has only been reported on a few occasions in Egypt, Kenya, Somalia, the United Arab Emirates, Pakistan, India, and Australia (Kinne et al. 2006). In addition to M. tuberculosis and M. bovis, a range of non-tuberculous mycobacteria was isolated from lesions resembling those caused by M. bovis in camels (Kinne et al. 2006). Generally, lesions occur in the lungs, bronchial and mediastinal lymph nodes, pleura, and liver (Mason 1917; Elmossalami et al. 1971). In tuberculous carcasses about 87.8% contained single lesions. Of the lesions in 91 camels detected postmortally during post-slaughter examination, about 50% had lesions in the lungs in which they were located predominantly in the apical and cardiac lobes, while lesions in the lymph nodes were most commonly present in the mesenteric nodes and less often in the mediastinal nodes. Another study reported that 57.5% of the lesions occurred in the thoracic lymph nodes and the lungs, 27.2% in the lymph nodes of the head, and 15% in the mesenteric lymph nodes (Beyi et al.
2014). Of the camels from which mycobacteria were isolated in one study, only two contained M. bovis, while the rest were infected with mycobacteria other than the M. tuberculosis complex (MTC; Mamo et al. 2011). The characteristic histopathological features of tuberculosis in camels resemble those seen in other animals, including the presence of acid-fast, rod-shaped bacteria (Kinne et al. 2006).
Sheep Sheep are considered, perhaps erroneously, not to be important in the epidemiology of BTB, and they are considered to be less susceptible to infection with M. bovis than both cattle and goats. However, cases of BTB have been diagnosed in New Zealand and Spain, and in a number of African countries (Davidson et al. 1981; Munoz-Mendoza et al. 2016). The lesions caused by M. bovis in sheep closely resemble those in cattle and vary from isolated mineralized granulomas in the lymph nodes to extensive, soft, caseous granulomas in the dorsal areas of the lung and some of its regional lymph nodes (Davidson et al. 1981). The lesions vary in size from disease has been detected in Africa, all were first diagnosed with BTB during postmortal examination, and this situation is expected to remain in the future. What is of importance in this respect is that there is a marked intra- and interspecific variation in the appearance of the lesions and in their distribution in the various species. The lesions in some species closely resemble those in cattle, but in others (particularly lions, leopards, and hyenas), they are distinctly different and bear no resemblance to the tuberculous granulomas generally associated with mycobacterial infections.
The pathology of the disease is inadequately described in all the African wildlife species in which BTB has been diagnosed. However, the differences in the appearance and distribution of the lesions in predators, antelopes, omnivores, and primates give an indication of the variation that can be expected, and investigators should not be misled by what has been described for long, as the “typical” tuberculous granuloma when investigating the presence of BTB in wildlife species. A further complication of detecting wildlife infected with M. bovis is the occurrence of BTB in those species that are not often encountered because of their small size and nocturnal habits, such as rats (Rattus norvegicus) (Little et al. 1982) and hedgehogs (Erinaceus europaeus) (Lugton et al. 1995) that have been found to be infected with M. bovis. Others have no visible lesions, and any one of them may play a role in the dissemination of the infection in ecosystems (Gavier-Widen et al. 2009).
The description of the lesions in the following species serves as an example of the diversity of lesions that may be encountered in African species of wildlife, and it is presented in some detail to guide further investigation of the disease in wildlife on the continent, and in the different African countries where they may occur.
African Buffaloes In African buffaloes, the lesions often resemble those seen in cattle in which they are characterized by containing a fibrous capsule, marked caseation, and some calcification. The various early developmental stages of the granulomas can similarly be classified as they are in cattle (Wangoo et al. 2005; Laisse et al. 2011). In buffaloes, some of the lesions are atypical and have a neoplastic, fibro-lardaceous appearance (Fig. 9.9), some contain branching interdigitating areas of caseous necrosis lacking calcification and a detectable capsule (Fig. 9.10), and others appear as well-encapsulated aggregates of an inspissated caseous necrotic debris (Fig. 9.11) (Guilbride et al. 1963; Woodford 1982). Pus may be present in some of the lesions in buffaloes, and it is invariably of a lighter color than that seen in tuberculous lesions in cattle. The caseous necrotic centers of the granulomas may become liquefied both in the lungs (Fig. 9.12) and in the lymph nodes. In advanced cases of the disease, the carcasses are cachectic.
The distribution of the lesions is quite consistent in that in most buffaloes with tuberculosis, lesions occur in the lungs (50% of cases had only pulmonary lesions), while in the largest proportion of infected buffaloes, lesions occurred in the bronchial and/or mediastinal lymph nodes. Rarely, lesions occurred exclusively in the retropharyngeal and cervical lymph nodes (Woodford 1982) or in the palatine
Fig. 9.9 Buffalo:
Iardaceous appearance of the granulomatous lesion seen on the cut surface of a mediastinal lymph node. Note the absence of necrosis and a distinct capsule
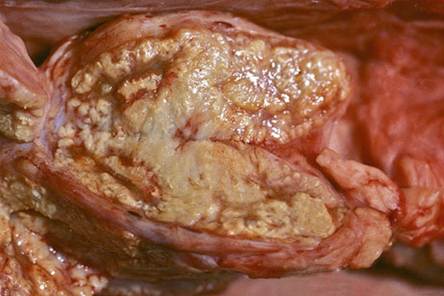
Fig. 9.10 Focal lesion in a lymph node of a buffalo manifesting interdigitating areas of necrosis, branching early fibrosis, and the lack of a distinct capsule (formalin- fixed specimen)
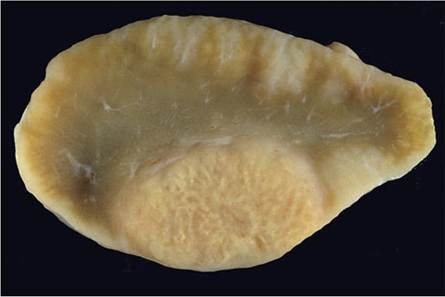
Fig. 9.11 Enlarged and encapsulated, inspissated, caseo-necrotic exudate in the mediastinal node of a buffalo
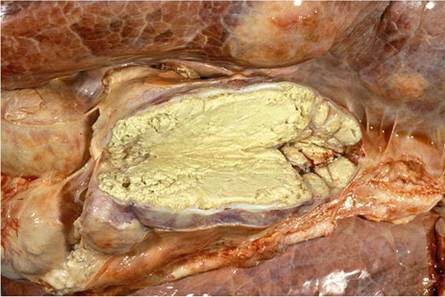
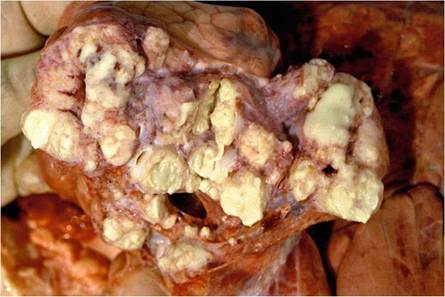
Fig. 9.12 Liquefied exudate in a multifocal pyogranulomatous pneumonia in an African buffalo
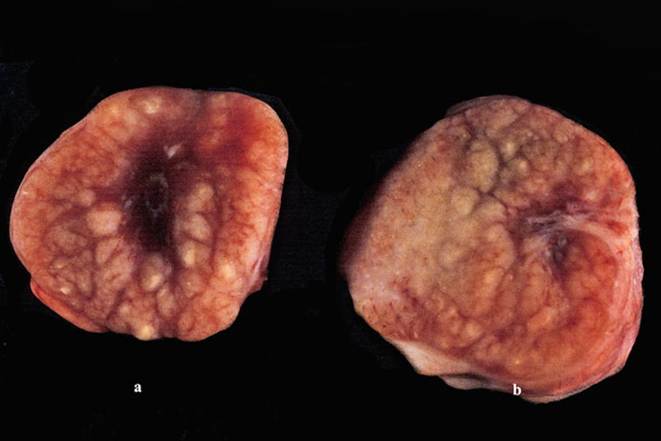
Fig. 9.13 Palatine tonsils. (a) Normal tonsil containing a few crypt abscesses. (b) This tonsil contains a poorly defined lesion partially effacing the normal structure of the tonsil. The granuloma blends into the normal tissue and is not encapsulated
tonsils. Lesions in the tonsils may be difficult to detect, and they are most easily located by palpating the organ as they cause an increased consistency of the affected parts of the tonsils. The granulomas in the tonsils are usually not encapsulated, and the reaction extends imperceptibly into the surrounding normal tissue and sometimes into the adjacent skeletal muscle (Fig. 9.13). On cursory examination, they may be confused with cysts and crypt abscesses that commonly occur in the tonsils in buffaloes (Kriek et al. 1994).
Pulmonary lesions may occur throughout the lungs, but they are most often found in the dorso-caudal aspects of the caudal lobes; often, only single lesions are detected, and they may be quite small and are then most easily detected by thorough palpation of the pulmonary parenchyma. The lesions vary in appearance from small almost undetectable, encapsulated, discrete, fibro-caseous granulomas to numerous coalescing foci (Fig. 9.14) of up to 30 cm in diameter. In many instances, they are distributed multifocally in the caudal portion of the lung where they may displace large portions of the lungs (Fig. 9.15), sometimes to the extent that more than 60% of the functional tissue has been destroyed. Lesions in the lung too may have a smooth lardaceous appearance (Fig. 9.16) and may vary in color; sometimes they may have an orangey appearance dependent on the characteristics of the strain of M. bovis that infected the animal. The granulomas commonly undergo limited to extensive caseous necrosis, and the necrotic exudate may become partially liquefied (Keet et al. 1994). Other organs and tissues are rarely affected, but when they are, the lesions
Fig. 9.14 Multifocal granulomatous pneumonia with a lobular distribution
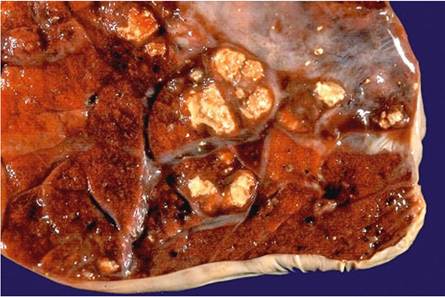
Fig. 9.15 Buffalo lung with an extensive lobar distribution of the lesions. Note the retention of the lobular structure
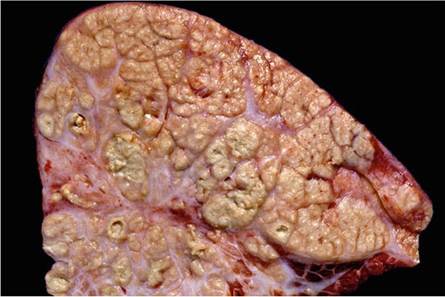
Fig. 9.16 Focal, locally extensive, lardaceous, granulomatous lesion in the lung. Note the orangey tinge of the affected tissue and the lack of the usual features associated with lesions caused by M. bovis
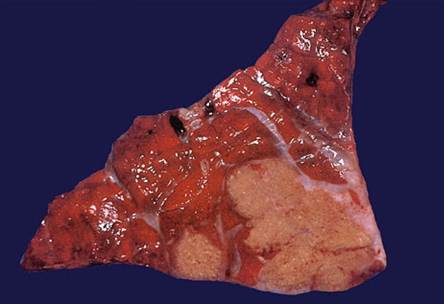
Fig. 9.17 Granulomatous enteritis causing slight, irregular, linear, elevation of the mucosa, containing an inspissated, partially calcified content when cut into
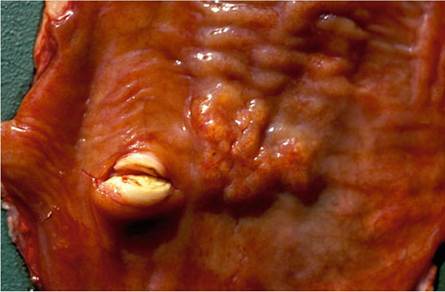
have the characteristic appearance of those seen in lymph nodes, the palatine tonsils, and the lungs. Elaeophora sagitta also causes localized parasitic granulomas in the dorso-caudal portions of the lungs, that are easily confused with those of BTB, thus emphasizing the need to confirm the cause of the lesion histopathologically.
Intestinal lesions are rarely seen in buffaloes, except in advanced cases in which they develop following expectoration and swallowing exudate containing large numbers of bacteria from advanced lesions in the lungs. The intestinal lesions are then more commonly seen in the distal portion of the small intestine, and at the ileocecal junction. Here they have a linear configuration and are mostly located within the mucosa and submucosa where their presence causes the development of small irregular ulcers (Fig. 9.17). Where these lesions occurred, the draining mesenteric nodes were always affected (Kriek et al. 1994).
Animals with generalized tuberculosis also manifested a granulomatous peritonitis, while a granulomatous pleuritis and pericarditis occurred in a quarter of the buffaloes with generalized BTB. Individual buffaloes rarely contain lesions only in
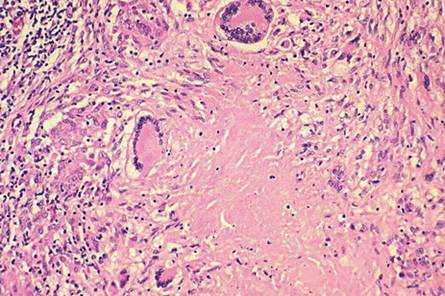
Fig. 9.18 Histopathological characteristics of a typical tuberculous granuloma. Note the central caseous necrosis and the presence of a number of Langhans giant cells, epithelioid cells, loosely arranged lymphocytes, and early fibrosis at the periphery of the lesion
the liver, spleen, or uterus and in the gastro-hepatic nodes. Peripheral lymph nodes are rarely affected but they do develop lesions in advanced cases of the disease.
The histopathological appearance of the lesions in African buffaloes differs little from that seen in cattle, the inflammatory reaction being characterized by a granulomatous reaction, the presence of epithelioid cells, Langhans giant cells, central caseous necrosis with some calcification, and a pronounced peripheral fibrous capsule (Fig. 9.18). Not all lesions are encapsulated. In some of the lesions, masses of neutrophils are present in the center of the necrotic portions, and these undergo liquefaction as a consequence of this inflammatory reaction. The numbers of acidfast bacteria seen in sections differ substantially; in some instances, numerous bacteria are scattered throughout the inflammatory reaction, while in others they are very sparse, and in a significant percentage of the cases, acid-fast organisms are not detectable (Woodford 1982).
Greater Kudus The lesions in the different organs are well demarcated, encapsulated, firm, and irregular, and have a caseous necrotic and granulomatous appearance on cut surface. In less severely affected animals, lesions are predominantly present in the lymph nodes of the head but without sufficient enlargement to cause visible swelling below the ears or in the parotid area. The nodes of the neck are less often involved, and lesions occur inconsistently in different lymph nodes, including the caudal cervical node. Multifocal, granulomatous, mediastinal lymphadenitis, and lesions in the lungs that are then limited to a few caseo-necrotic granulomas scattered throughout the pulmonary parenchyma, may occur in these early cases (Thorburn and Thomas 1940).
The irregular, and often prominent, swelling in the parotid area is a characteristic feature of the disease in some kudus with tuberculosis. When cut into, they contain cavities filled with a watery, turbid, whitish, purulent exudate enveloped by a thin,
Fig. 9.19 The typical fluid purulent appearance of the exudate surrounding exiting structures on the side of the neck and in the parotid region of a kudu
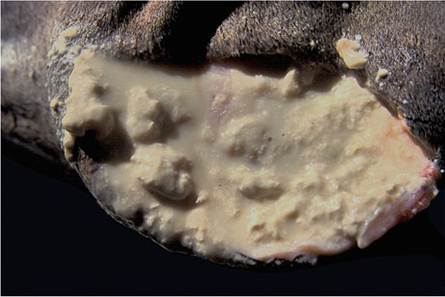
well-vascularized capsule (Fig. 9.19). The pressure exerted by the large volumes of accumulated exudate causes it to extend between the anatomic structures of the neck (Weber and Van Hooven 1992). Ventral to the ears (where the parotid salivary gland and lymph nodes are situated), large, poorly encapsulated, caseo-necrotic masses replace the normal tissue. In many instances, these lesions drain through a cutaneous sinus (Thorburn and Thomas 1940).
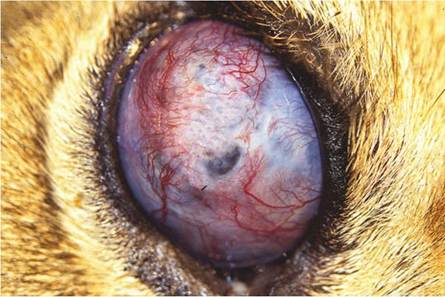
In advanced cases, the disease becomes generalized. All the lymph nodes of the head, neck, thoracic cavity, as well as some of the caudal cervical, mesenteric, and the hepatic nodes, are often involved. The bronchial nodes are usually markedly enlarged (up to 10 cm in diameter), and their normal structure is completely replaced by a severe, granulomatous inflammatory reaction that also contains large masses of caseo-necrotic debris. Lymphatics draining into a lymph node are often affected by a multifocal, granulomatous lymphangitis, the individual granulomas in the lymphatics being up to 2 mm in diameter. Corneal opacity and a fibrinous exudate in the anterior chamber of both eyes may be present indicating spread to the ocular structures in some of the advanced cases (Keet et al. 2001).
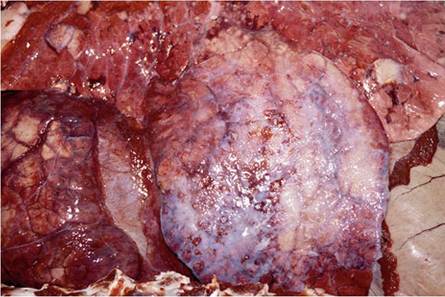
The lungs in advanced cases are severely affected, and the physical appearance of the granulomas varies according to their locality. Those in the cranial lobes are larger and more numerous than in the caudal lobes and replace most of the normal parenchyma forming extensive areas of consolidation. The granulomas in the cranial lobes are clustered, even-sized (30-40 mm in diameter), ovoid, oblong or triangular, and well encapsulated (Fig. 9.20). They contain a smooth and inspissated caseo- necrotic exudate that may be partially liquefied and poorly calcified. Similar granulomas occur in the caudal portions of the lungs, but as a few isolated masses scattered randomly throughout the lobes. In addition, miliary tubercles appearing as pearly areas of granulomatous inflammation of about 3-5 mm in diameter can be scattered throughout the caudal lobes. These small granulomas tend to be clustered, thereby imparting a mulberry-like appearance to the lesion. A diffuse granulomatous pleuritis that predominantly involves the affected portions of the cranial lobes, results in fibrous adhesions to the parietal pleura (Fig. 9.21) (Keet et al. 2001).
Fig. 9.20 Lung of a kudu manifesting the typical, well-demarcated, and often angular, granulomatous lesions seen in this species
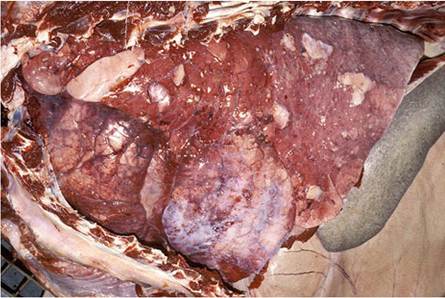
Fig. 9.21 Diffuse, chronic pleuritis with extensive fibrosis on the apical lobe of the lung in a kudu
Histologically, the lesions are encapsulated and contain extensive areas of caseous necrotic debris surrounded by a typical granulomatous inflammatory reaction containing numerous epithelioid cells, Langhans giant cells, and lymphocytes. The lesions are further characterized by the presence of large numbers of neutrophils particularly in the caseous necrotic debris. Even very early lesions contain aggregates of neutrophils. Few acid-fast bacteria are present in the cytoplasm of Langhans giant cells and in the exudate (Keet et al. 2001).
African Lions The lesions caused by M. bovis in lions are substantially different in their macroscopical and histological appearance compared to the lesions seen in other wildlife species in which this infection occurs (Keet et al. 1998). Histologically, homogenous infiltrates of macrophages, the lack of necrosis, calcification, and the absence of Langhans giant cells characterize the lesions. Normally mycobacteria are absent from these lesions.
In the lungs, the inflammatory reaction is initially limited to the interstitial tissue, while a mixed macrophage and neutrophil exudate may be present in the
Fig. 9.22 Lion lung with two defined, focal, raised foci characteristic of the lesions in the early stages of the disease
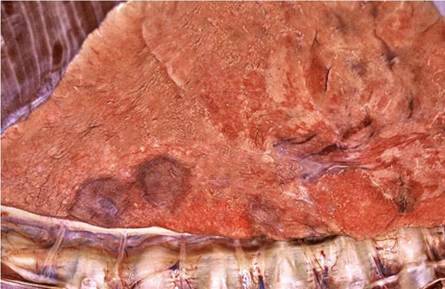
alveoli. The areas of granulomatous inflammation are interspersed with foci of an acute inflammatory reaction containing fibrin, a neutrophil exudate, and a marked inflammatory edema. The alveolar walls are thickened and infiltrated by a mixed inflammatory exudate consisting of macrophages, fibroblasts, and proliferating type 2 pneumocytes. Marked bronchiectasis is seen within which there is a copious, mucopurulent, inflammatory exudate in which large numbers of mycobacteria can be seen following staining with the Ziehl-Neelsen (ZN) staining technique.
The initial macroscopical lesions in the lungs are nondescript, raised foci of a few centimeters in diameter that bulge slightly above the pleural surface and fade into the surrounding tissue, and are most easily detected following collapse of the lungs after the thoracic cavity is opened (Fig. 9.22). In more advanced cases, these lesions become confluent and affect large portions of the lung (Fig. 9.23). The lesions, when cut into, are ill defined and contain a semifluid mucopurulent exudate (Fig. 9.24) that originates from the dilated portions of the bronchi that are the consequences of bronchiectasis (Fig. 9.25). The parenchyma surrounding the dilated bronchi contains an inconspicuous granulomatous inflammatory reaction seen as a collar of pale, tan-colored tissue that blends into the surrounding normal parenchyma.
The lymph nodes of these animals may be enlarged, but it is not possible to detect lesions macroscopically because of the nature of the inflammatory reaction that lacks the normal features of a mycobacterial infection. In many instances, the lymph nodes may also contain large cystic spaces that extend throughout the node, but these lesions too, contrary to some anecdotal opinions, have no diagnostic significance when dealing with tuberculosis.
Lions suffering from BTB are emaciated in the terminal stage of the disease and often have large cutaneous granulomas and arthritis caused by the infection. The elbow joints may be substantially enlarged by a chronic arthritis, and contain numerous fibrin globules as part of the exudate (Fig. 9.26). Hypertrophic osteopathy may be seen in the long bones of some of the infected animals in association with joint lesions. Occasionally, panophthalmitis may be seen as a consequence of the infection (Fig. 9.27). In many of the terminal cases of BTB, renal amyloidosis may
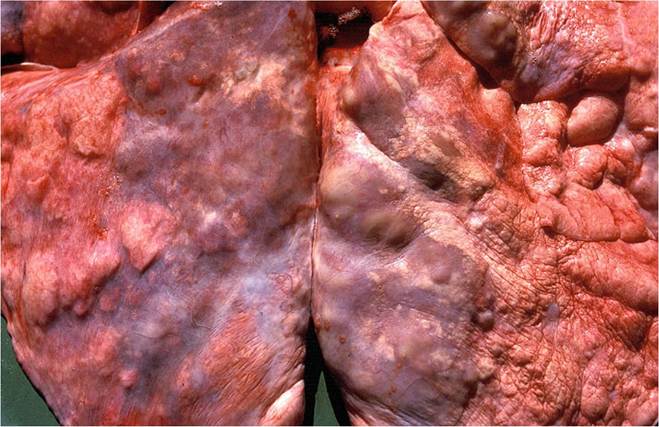
Fig. 9.23 The atypical lesions in the lung of a lion in the advanced stage of the disease resembling and extensive interstitial pneumonia
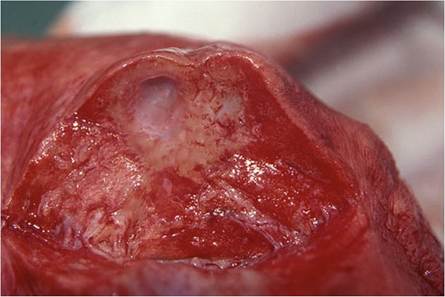
Fig. 9.24 Cavities caused by extensive bronchiectasis in the lung of a lion. These bronchi contain copious amounts of a mucopurulent exudate
be detected. This lesion may or may not be directly related to the mycobacterial infection, since it is also seen in free-ranging lions that do not suffer from the disease.
Lesions in leopards and hyenas are very similar to those seen in lions. In cheetahs, the appearance of the lesions in a single animal was recorded, and they differed markedly from those seen in lions. Macroscopically, the only lesions occurred in the lungs as extensive areas of necrosis in a multifocal to confluent granulomatous
Fig. 9.25 Cavity caused by bronchiectasis in a lion’s lung from which the exudate has been washed off to reveal the dilated bronchus
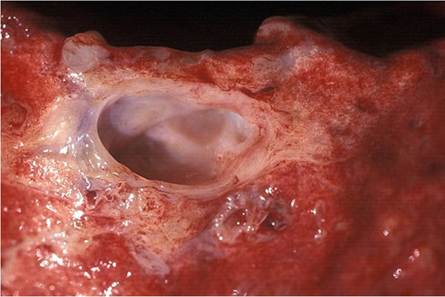
Fig. 9.26 Severe chronic, deforming arthritis caused by an M. bovis infection in a lion
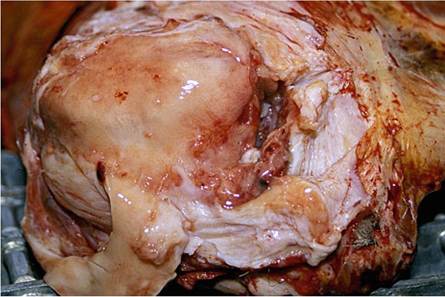
Fig. 9.27 Severe chronic granulomatous panophthalmitis in a lion
pneumonia containing a fluid, purulent exudate. Histologically, the lesions were characterized by the presence of multiple aggregates of neutrophils, occasional foci of coagulative necrosis, limited fibrin exudation, and limited calcification of a poorly encapsulated expanding granulomatous inflammatory reaction. The alveoli surrounding the inflammatory response contained macrophages and epithelioid cells, and the associated terminal bronchioles were plugged with a necrotic exudate containing many neutrophils. Scattered acid-fast bacteria occurred in the exudate and intra-cytoplasmically in macrophages at the periphery of the lesions (Keet et al. 1996).
Chacma Baboon The macroscopic lesions are typical tuberculous granulomas that may occur throughout the body. The distribution of the lesions suggests that there is early postprimary embolic spread following the initial infection. Lesions appear consistently in the mesenteric lymph node, spleen, and lungs (Fig. 9.28). The lymph nodes of the head and neck are inconsistently affected, as are others such as the mammary, inguinal, and axillary lymph nodes. In individual cases, lesions may also be present in the liver and kidneys. Tuberculous granulomas may occur in some of the vertebra.
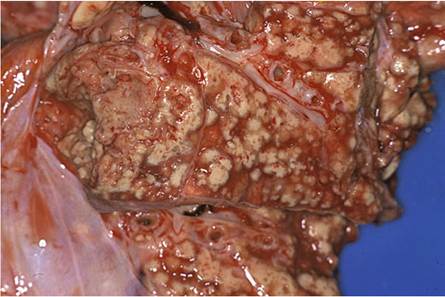
The lesions are commonly well-circumscribed with a multifocal to confluent distribution. Their consistency varies from solid in those with fibrosis, to soft when the content liquefies. The initial lesions in the lungs have a miliary distribution reflecting a typical embolic origin. In time, they become larger and confluent and the exudate then may become liquefied, and cavities in the lung tissue may develop (Fig. 9.29).
Histologically, the lesions are characterized by areas of central caseation, containing aggregates of necrotic neutrophils within a multifocal to coalescing granulomatous pneumonia. Infiltrates of macrophages, epithelioid cells, and Langhans giant cells, lymphocytes, and plasma cells surround the necrotic centers of the granulomas. The lesions are poorly encapsulated, and a miliary spread can be seen. Liquefied exudate within granulomas may drain into adjacent bronchioles and
Fig. 9.28 Extensive, miliary, granulomatous pneumonia in a baboon following late, postprimary dissemination of the infection
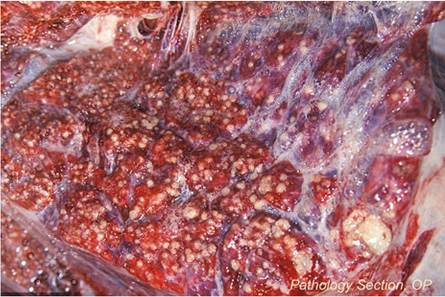
Fig. 9.29 Multifocal to confluent, granulomatous pneumonia in a baboon. Note the liquefaction of the exudate on the left, and collapse of the tissue resulting in cavitation
bronchi in which the mucosa contained a multifocal caseous granulomatous inflammation causing ulceration in areas (Keet et al. 1996). Granulomatous lesions may also be present in the liver, spleen, and lymph nodes. Many acid-fast bacteria occur within the lesions (Keet et al. 2000).
Warthogs Woodford (1982) described the lesions in warthogs infected with M. bovis. They were seen as calcified abscesses in the submaxillary lymph nodes and lungs. The lesions in the lungs may be extensive and consist of a caseo-calcific consolidation of the entire lung due to coalescing masses of small granulomas with an embolic distribution. In generalized cases, lesions may also be seen on serosal surfaces, and there may be dissemination to many lymph nodes. Both M. bovis and atypical mycobacteria may be isolated from lymph nodes with lesions.
Black Rhinoceros The lesions described in one case were limited and presented as multifocal, firm and irregular granulomas varying in diameter from 1 to 6 cm in the dorso-cranial portions of the lung lobes. They were encapsulated and contained a creamy necro-caseous exudate in which numerous acid-fast mycobacteria were detected in smears of the exudate (Miller et al. 2017).
Based on the appearance of the lesions in the various species, it must be clear that one should expect a substantial variation in their macroscopical appearance and distribution. It should be easy to make a presumptive diagnosis in advanced cases and in those with single or only a few large lesions. The challenge lies in detecting the lesions when they are single, small, or not visible and when they occur in sites in which they normally are not expected. The likelihood of missing the diagnosis with a cursory examination emphasizes the need to do a complete necropsy, should the intention be to use it for surveillance purposes. The implications of ignoring lesions in wildlife species and considering them to be “just abscesses,” as has been done in the past, may have important implications as the disease then spreads imperceptibly throughout the ecosystem for many years before it is finally diagnosed.
9.6.4 Detection of Lesions Caused by M. bovis in Livestock in Abattoirs
There are different reasons for detecting lesions caused by M. bovis infection in domestic and wild animals. These include abattoir examination for the soundness of meat for human consumption, surveillance purposes for the control of BTB, and research. A single procedure for reliably detecting lesions that would satisfy the demands of each of these activities does not exist. Each of the different activities requires a different level of accuracy, and thus more or less attention to detail when doing the examinations. If you do not want to do what is required, or do not have the inclination to do an appropriate examination fit for the purpose of the investigation, do not bother to do it, because you will be wasting your time. Different protocols have been designed for each of these activities, and they will be dealt with in the following sections.
Meat Inspection in Abattoirs Routine abattoir inspection is based on procedures legislated by animal health authorities and/or the quarantine division of the Ministry of Agriculture in individual countries (Murray 1986; Kaneene et al. 2006; Awah- Ndukum et al. 2012; Koro et al. 2013). Inspection is intended to be conducted by trained meat inspectors with the main objective of ensuring that meat and organs are fit for human consumption. This includes detecting the presence of BTB that will, depending on the extent and distribution of the lesions, prompt rejection of affected organs or tissues, or result in total condemnation of the carcass for human consumption (Etter et al. 2006). The procedure varies somewhat between countries. In some, a specific set of lymph nodes must be incised and the cut surface inspected for the presence of lesions. In others, the nodes are merely palpated and assessed visually, a procedure that may detect 16% fewer lesions, and is thus substantially less Se (Corner 1994). Further inspection involves visual examination and palpation of organs, such as the liver and kidneys, and palpation and in situ incision of the tracheobronchial, mediastinal, and caudal cervical lymph nodes and the lungs. Other lymph nodes and organs are incised if lesions are detected in one of these tissues. In some countries, the mesenteric lymph nodes and lymph nodes of the head (Awah- Ndukum et al. 2010) and supra-mammary lymph nodes (Ngandolo et al. 2009) must also be routinely inspected.
By following the protocol for routine meat inspection, 43% of tuberculous animals may be missed (Corner 1994), while Bekele and Belay (2011) and Aylate et al. (2013), respectively, found that as many as 90.5 and 87% of cattle with tuberculous lesions, were missed. Stark et al. (2014) similarly found that the success of meat inspection in general in detecting lesions is highly correlated with the presence of clinical and/or pathological signs in affected animals and that early or subclinical cases are likely to be missed at slaughter. The impact of the use of visual- only inspection instead of incising the lymph nodes and tissues was negligible for most notifiable diseases and conditions with the exception of detectable cases of tuberculosis. In addition to the specific methodology used, the success in detecting lesions during the course of abattoir inspection will also depend on the competency of the examiner, and the time spent on examining the animal. One investigation reported that the mean time spent to inspect a carcass (1.2 ± 0.4 min) by the routine method was virtually five times shorter than the detailed method (5.8 ± 1.9 min) (Bekele and Belay 2011), hence the difference in the successful detection of lesions when they are present.
Meat inspection at abattoirs is the main test method used in the final stages of eradication programs and for documenting freedom from BTB in developed countries from which the disease in cattle has been eradicated. The lack of the sensitivity of using routine meat inspection as a means of surveillance in the advanced stages of eradication schemes, although it may be much less expensive than sustaining the use of TSTs, is thus not without risk. This is particularly applicable when the issue of not being able to detect cases with non-visible lesions with current technology, is taken into consideration (Gavier-Widen et al. 2009).
Detailed Postmortem Inspection Under certain circumstances, for instance, for research purposes and the reliable detection of lesions in positive reactors, because of the low sensitivity of routine meat inspection to detect BTB, it is necessary to increase the sensitivity of postmortal detection of BTB-positive carcasses. The sensitivity of the investigation can be improved substantially by simply examining six pairs of lymph nodes, a process that is sufficient to detect 95% of cattle with macroscopic lesions (Corner et al. 1990). The specificity of macroscopic examination by the parallel use of histopathology can also enhance the Se of the investigation to 94-95% (Rohonczy et al. 1996).
Because of a long list of tissues, organs, and anatomical sites affected by BTB, the results obtained vary greatly in scope from study to study (Corner 1994). Commonly, the following lymph nodes are sliced at a thickness of 2 mm: parotid, retropharyngeal, mediastinal, tracheobronchial, mesenteric, submaxillary, iliac, precrural, prescapular, supra-mammary, inguinal, ischiatic, portal, and sternal. Organs including the lungs, liver, kidneys, mammary gland, intestines, heart, and meninges are thoroughly examined visually and by palpation and are incised at thicknesses varying from 2 to 20 mm (Asseged et al. 2004; Biffa et al. 2010). The cut surfaces are examined under a bright light source for the presence of TB-like lesions. Sectioning and the detection of small lesions are easier after overnight refrigeration of the lymph nodes. To increase the sensitivity of the detailed examination, aseptic specimens from all lymph nodes should be submitted for culture, according to the protocol used (Corner 1994).
Meat Inspection in Africa With the exception of the situation in a few countries, meat inspection at abattoirs in Africa is in a parlous state (Bekele and Belay 2011). The purpose of meat inspection is to focus on the removal of grossly abnormal meat, organs, and related products from the food chain, thus preventing the distribution of infected products that could cause disease in humans. It should also assist in the detection of specific diseases to allow tracing-back infections to their source for control purposes (Biadglegne et al. 2013; Habarugira et al. 2014). Of these, the removal of grossly abnormal products for human consumption is probably the easiest to accomplish and is overemphasized in Africa.
Meat inspection protocols currently utilized in abattoirs in Africa are generally insufficient to detect the majority of visible TB lesions. Factors contributing to the low sensitivity include the lack of adequately trained personnel, heavy workload, poor physical facilities with inadequate lighting, and the lack of diligence of the inspectors (Awah-Ndukum et al. 2010; Biffa et al. 2010). Furthermore, as the examination is performed under variable conditions, inconsistencies of interpretation are difficult to avoid. Corruption, to limit the impact of the loss incurred by condemned carcasses, has also been highlighted as a limiting factor in applying the existing regulations (Sulieman and Hamid 2002).
Standardization of abattoir inspection protocols (in line with international sanitary requirements), enhanced training, quality control measure to assess the standard of meat inspection, and raising public awareness are essential and cost-effective measures that should be implemented to improve the situation (Biffa et al. 2010) and to increase the accuracy of diagnosing BTB in abattoirs.
The specificity of meat inspection as a means to detect BTB in Africa is also affected by a plethora of non-specific TST reactions caused by environmental mycobacteria and the presence of lesions caused by parasitic infections and bacterial or mycotic granulomas and abscesses caused by pyogenic bacteria, which may be macroscopically indistinguishable from tuberculous granulomas (Liebana et al.
2008). A number of examples are quoted to demonstrate the extent of the problem. A study in Mali revealed that 72% of tuberculous lesions detected during routine meat inspection were due to pathogens other than M. bovis (Maas et al. 2013). In another report, only 11.2% of BTB-like lesions in Tanzania were positive for M. bovis (Mwakapuja et al. 2013). Various pathogens including the pulmonary form of nocardiosis (in Sudan) (Awad 1962), granulomas caused by parasites such as liver flukes, actinomycotic granulomas (Berrada 1993), and fungi (Kuria and Gathogo 2013) may be confused with the lesions of BTB. In addition, NTMs (up to 44.8% in certain studies) cause similar lesions to those caused by M. bovis that may result in incorrect diagnoses in the absence of the ability to confirm the diagnosis, at least by histopathology, and, preferably by culture (Berg et al. 2009; Sahraoui et al. 2009; Biffa et al. 2010).
The limitations of using abattoir diagnostic data as a means of surveillance for BTB in Africa must be kept in mind by regulatory authorities and policy makers. Using these data also have an effect on the time it takes to detect an infection in a herd as the estimated median time for detection by abattoir surveillance of a herd after the disease was introduced was 5.75 years compared to 1.99 years when a TST was used as a routine diagnostic procedure (Fischer et al. 2005). However, in Africa, as monitoring of BTB using bacteriology and TSTs is time-consuming and prohibitively expensive, and the laboratories are ill-equipped, it is inevitable that data obtained from meat inspection will be the only way in which some useful information about the extent of BTB in the continent’s livestock population can be gathered. To do this, the process will have to be exploited to its full potential, always keeping in mind that the process is flawed (Atiadeve et al. 2014).
9.7
More on the topic Postmortal Examination (PME):
- General Physical Examination
- Examination by Age
- Examination
- Physical Examination
- Gross Examination
- Sensory Examination
- History and Examination
- Examination of the External Ear Canal
- Elite-formation: the Army, the Lineage and the Examination System
- Selecting the appropriateimaging method, correctly applying the technique selected, and accurately interpreting the examination are the key steps in imaging ear disorders in dogs and cats.
- Ophthalmic examination is an integral part of general clinical evaluation, not only to identify local disease but also to suspect or detect many systemic abnormalities. Eye is also considered as a window to visualize intracranial pathology.
- Careful examination of a clean, dry ear canal in a dog or cat with otitis externa may reveal many conditions that affect the ear canal.
- The physiatric history and examination of a child require a blend of medical diagnostic skills to establish or confirm the diagnosis as well as a knowledge of child development and behavior to evaluate functional assets and difficulties for the intervention phase of rehabilitation.
- Evaluation of a child presenting in the health care involves four key components, often referred with acronym HELP, i.e. History, Examination, Logical deduction and Plan of investigation. Present chapter outlines broad guidelines regarding history-taking, the first step in evaluation process.
- Chapter Eight Seung Ho Bang, Oded Borowski, Kook Young Yoon, and Yuval Goren Local Production and Domestic Ritual Use of Small Rectangular Incense Altars: A Petrographic Provenience Analysis and Examination of Craftsmanship of the Tell Halif Incense Altars
- Control of BTB in Cameroon
- ABDOMINAL MASS/LUMP
- CLINICAL EVALUATION OF GIT DISEASE
- Neuromuscular System
- Chest Pain