The Immune System
As stated above, the immune system can be subdivided into two parts, cellular immunity and antibodies, although within the animal the two systems will work very much in conjunction with one another to counteract disease.
Although I shall be dealing with the immune response to disease-causing organisms, the reader should appreciate that an identical immune reaction is evoked against any material which the animal recognises as being foreign to its system. This is very important in the human fields of allergy and organ transplant rejection. Any material which the animal recognises as foreign is called an antigen, and antigens evoke both a cellular and an antibody response. Immunity consists of both innate and induced components.Innate mechanisms
Cellular response The most common cells involved with innate immunity are neutrophils and macrophages. Both engulf and destroy invading infectious agents by a process known as phagocytosis. Neutrophils and macrophages are therefore sometimes collectively known as phagocytic cells, or simply as phagocytes.
The process of phagocytosis is shown diagrammatically in Figure 1.3. Neutrophils are commonly present in blood, whereas macrophages can be found in milk and other secretions. Macrophages are a type of ‘bobby on the beat’. When they see something they don’t like, they ‘arrest it’ (engulf it) and at the same time send out a signal which mobilises a ‘rapid reaction force’ of highly active neutrophils. The neutrophils then continue with the process of phagocytosis and destruction.
Macrophages (and cancer cells) produce matrix metalloproteinases (MMPs). These enzymes dissolve body tissue and allow the macrophages to pass between the body cells in their search for foreign invaders. MMPs also allow cancer cells to penetrate. Another important function of macrophages is that they can also hold the invading antigen in a very specific manner.
They then present it to the lymphocytes (another form of white blood cell) of the induced immunity system, to ensure that the lymphocytes will recognise it in the future.Humoral response The humoral part of the innate immune system consists of proteins such as complement, interferon and lysozyme. Complement coats the outside of invading agents, in a process known as opsonisation. This makes the invading organism more easily engulfed by macrophages. Interferon is best known for its effect against viruses, but it also has a role in neutralising toxins. Lysozyme is a form of ‘natural antibiotic’ and is highly active in killing bacteria.
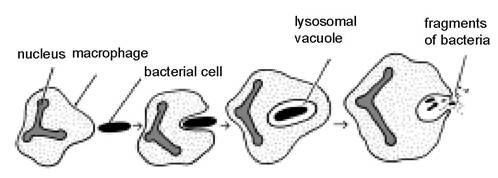
Figure 1.3. Phagocytosis, the process by which an animal cell recognises and then engulfs and destroys foreign substances such as bacteria and viruses.
Induced mechanisms
This also has cellular and humoral components, but each cell and each antibody is highly specific to the invading agent. The induced immune system differs from the innate immune system, in that the induced system must have had previous exposure to an antigen in order to be effective.
Cellular response The cellular response of the induced immune system is often referred to as cell mediated immunity. The major cells are lymphocytes.
The body's defences against infection are
Physical barriers
Chemical barriers
The immune system
- antibodies (humoral mechanisms) and cells
- innate and induced systems
There is a range of different lymphocytes. They all produce antibodies, but some may have additional functions and means of recognising and destroying invading antigens. T lymphocytes, including killer T cells, recognise foreign cells, for example cancer cells and tissue transplants in man. Cells which are infected with virus will also be recognised and destroyed by the killer T lymphocytes. ‘Helper’T lymphocytes (also known as T4 cells) hold the invading antigen so that it can be recognised by B lymphocytes.
Each B lymphocyte (also known as a plasma cell) has thousands of recognition sites on its surface.To return to our police force analogy, macrophages and T lymphocytes hold invading antigens (‘suspects’) and present them to B lymphocytes (‘policemen’), thus enabling the suspects’ fingerprints to be taken. Each lymphocyte carries up to 100,000 different fingerprints. Considering there are millions of lymphocytes, this makes the total number of combinations almost infinite. Each recognition site is different for every individual invading antigen. When a lymphocyte meets up with its specific antigen, two things happen:
• First, that single lymphocyte multiplies rapidly, producing clones of other identical lymphocytes which are immediately able to recognise that specific invader in the future.
• Secondly, these lymphocyte clones then produce antibodies, namely specific proteins to neutralise the invader.
It is interesting to note the numbers of cells which are involved. In an adult dairy cow approximately 8% of its bodyweight is blood, that is 48 litres for a 600 kilo cow. An average cow has around 7000 white blood cells per millilitre of blood, approximately 35% of which are lymphocytes, which means there are 2450 lymphocytes per millilitre of blood, or 117,600,000 lymphocytes in total! This only counts the lymphocytes in the blood. Lymphocytes are continually able to move out through the walls of the blood vessels into the extracellular fluid space, across to lymph nodes and then back into the blood again, all the time looking for ‘foreign invaders'.
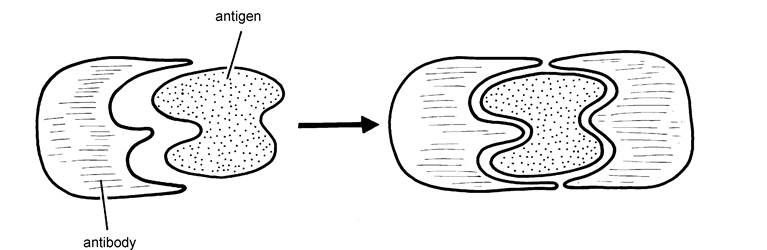
Figure 1.4. Antibodies are proteins. They work by fitting precisely into the shape of an invading antigen, thereby neutralising it.
Humoral response Antibodies are large protein molecules produced by lymphocytes to combine with, and hence neutralise, the invading agents. The most interesting feature of antibodies is that they are very specific.
Whereas the other defence mechanisms we have discussed so far will be effective against any bacteria, viruses or even dust, there has to be a separate and specific antibody for every type of infectious agent. Thus antibodies effective against one type of E. coli bacteria may not have any action against a slightly different strain of E. coli.Antibodies work by precisely fitting the shape of the invading antigen, for example, bacteria or virus. This is shown in Figure 1.4. The resultant complex neutralises the antigen, rendering it inactive and no longer capable of further invasion of body tissue. In addition, the antigen/antibody complex is more easily phagocytosed by macrophages. The ‘fit' of antibody to antigen needs to be precise to be effective. Some of the less effective vaccines may produce antibodies which are not exactly the correct shape. Although they may be able to ‘arrest' some of the invading infection, other ‘invaders' are able to break free and still cause damage. This is shown in Figure 1.5.
Antibodies are acquired by the animal in two separate ways known as active and passive. Active immunisation is the process whereby the animal produces its own antibodies following exposure to an antigen. The cow can also produce antibodies and supply them preformed to the calf during the first few hours of its life via the thick first milk called colostrum. Because the calf has not produced these antibodies itself, they are called passive and they provide immediate protection against infections present in the environment.
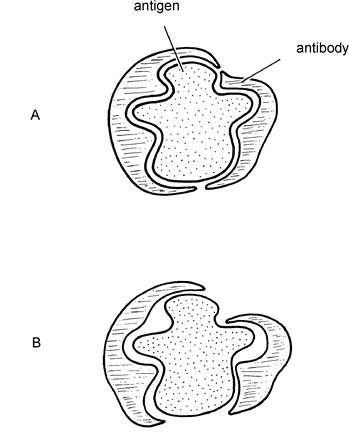
Figure 1.5. If the antibody is a poor fit (i.e. not totally specific) as in B, the antigen may break free and continue to cause damage to the host.
Before an animal can produce its own active antibody against a particular infection, it must have been exposed to that infection at some time in the past, recognised it as foreign (viz as an antigen) and stored the information in a type of memory, ready to produce antibodies to overcome subsequent challenges.
This initial exposure may be by vaccination, but it is much more likely to be the result of natural infection. A low dose of disease organisms which is not sufficient to cause visible symptoms will be quite adequate to stimulate antibody production and provide active immunity. This process is occurring throughout the animal’s life, and re-exposure to infections helps to boost immunity levels.Vaccines will be used when there is a risk of a heavy challenge from a specific infection, and especially if the animal has not had previous exposure to that infection. A vaccine consists of the infectious agent which has been altered in some way. When administered to the animal it stimulates the processes of recognition and antibody production, but it cannot cause disease. Vaccines may either be living, when only one dose may be required, or dead, when two doses will be needed at an interval of approximately four weeks. The presence of passive immunity, that is antibodies acquired from the mother, may prevent the calf from responding to the vaccine and this is why the instructions may state that animals under a certain age should not be vaccinated, or perhaps that if young animals are vaccinated, then an additional dose may be necessary at a later date. Passive immunity generally persists until the animal is two to three months old, depending on the amount of antibody received in the colostrum, and on the type of infection although there are exceptions to this. Vaccination of the calf should therefore be carried out at such an age that the period between passive and active protection is minimal, but not too early so that there is a risk of a poor vaccine ‘take’ due to persistence of passive colostral immunity.
Antibody titres
Antibodies are very specific: there is a group of antibodies for infection A, another group for infection B and so on. The level, or concentration, of antibody in the blood is referred to as the antibody titre. This may be expressed as
1:50 - blood can be diluted 50 times before the specific antibody can no longer be detected
1:100 - blood can be diluted 100 times before no antibody can be detected 1:1500 -blood can be diluted 1500 times before no antibody can be detected
Clearly the animal with a titre of 1:1500 has more antibodies to a particular disease than the animal with a titre of only 1:50.
The titre tells us nothing about the source of the antibody. It could be
• from colostrum - in which case the titre would slowly decline as the antibodies become worn out
• from a recent infection - in which case the titre would be rising, with the antibody-producing lymphocytes being in a production mode, having just been exposed to infection
• an old infection - in this case the titre would be slowly declining, unless there was a more recent exposure to the same infection, when the antibodies would start to rise again
Antibody titres are sometimes used to diagnose the cause of disease, for example calf pneumonia. If a blood sample is taken as soon as the calf is seen to be ill, then the antibody level to whatever is causing the disease is likely to be low (unless there is still some colostral antibody remaining). A second blood sample is taken two to three weeks later, by which time the amount of antibody to the infectious agent producing the high temperature should have increased considerably. The two blood samples are then tested for antibody levels to a range of possible infections, for example RSV, IBR and PI3 in the case of respiratory disease. The virus which shows a significant increase in antibody titre between initial infection and three weeks later is likely to be the cause of disease.
It is because of antibodies and other defence mechanisms that an animal can have bacteria living in it without succumbing to disease. The situation can rapidly change however if we mix groups of animals, for example calves from different sources. Calves from one farm may be carrying infection A and have antibodies to A. Calves from a second farm have infection B and the corresponding B antibodies. When the two groups are mixed, calves from A farm are exposed to infection B, but they only have antibodies to A. If the dose of B infection is large enough (for example if the groups were mixed and crowded into a poorly ventilated building), then B disease may occur in A calves before they are able to build up sufficient antibodies against B for protection. This is shown diagrammatically in Figure 1.6.
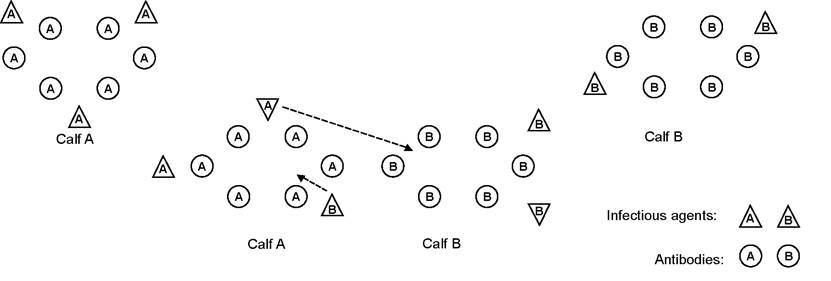
Figure 1.6. Specificity of antibodies. Calf A can exist in the presence of infection A because it has antibodies to A. Similarly calf B can exist with infection B. Problems occur when the calves are mixed. Calf A may be totally overwhelmed by infection B before it has had time to develop antibodies to it.
Stress and the Immune System
Environmental stress in cattle (and all farm animals) is a major problem, because it decreases the functional capacity of the immune system. Put another way, we have discussed the many remarkable ways in which an animal is able to counteract invasion by disease agents. However, if an animal is stressed, then these defence mechanisms simply do not work as effectively. Stress leads to the release of hormones such as adrenalin and cortisone, adrenalin preparing the animal to run away, cortisone specifically reducing the activity of its immune mechanisms. So what is stress? Examples of stress leading to a reduced immune response include:
• poor nutrition, including specific deficiencies of vitamins and minerals
• overcrowding, for example the lack of a loafing area. Animals are unable to move away from one another to find any ‘personal space’
• fear, for example young heifers introduced into a large dairy herd
• uncomfortable accommodation. Perhaps heifers which are not cubicle-trained lie outside on hard, wet concrete. Poorly ventilated cubicle buildings, with condensation dripping onto the cows’ backs, are also a stress
• rough and unsympathetic handling: driving cattle hard, using dogs or tractors, all produce fear
• excessive noise (this would be important for sows in farrowing houses)
• transport. Transport stress is interesting. Experimentally it has been shown to cause an increase in antibodies but a decrease in cytotoxic T cells, ie the lymphocytes which destroy virus-infected cells. This is probably one reason why animals succumb to virus infections after transport
• severe competition for food or water
• concurrent diseases, e.g. chronic lameness or rumen acidosis
• weather. Temperatures above 25°C have been shown to decrease the white blood cell count and therefore compromise the immune response
• calving. Both the cow and calf have a poor immune response for the first week after calving and should therefore be housed and managed to minimise further stress. Stress in heifers is also discussed in Chapter 9
Not only will the above environmental influences reduce the animal’s resistance to disease but perhaps equally as important, they can reduce the effectiveness of vaccines. In other words, a stressed animal will not respond as well to vaccination.
More on the topic The Immune System:
- Deadly Intentions: Apoptosis in the Peripheral Immune System
- Rheumatic disorders (connective tissue disorders or collagen vascular disorders') is a collective term to denote a large group of conditions with variable manifestations but two common characteristics: (a) acute or chronic inflammation of multiple target organs, specially musculoskeletal system and vasculitis, and (b) an underlying abnormal immune response, e.g. autoimmune etiology.
- Immune Thrombocytopenia
- Immune Response
- Immune systems, biochemical defenses, and symbionts can protect hosts against parasites
- CAUSES AND MECHANISMS OF IMMUNE ACTIVATION IN HIV INFECTION
- CONSEQUENCES OF IMMUNE ACTIVATION
- Specific immune responses to HIV
- IMMUNE THROMBOCYTOPENIC PURPURA
- ENVIRONMENTAL-MEDIATED AVAILABILITY OF VITAMIN D AND IMMUNE FUNCTION
- APPROPRIATE T CELL DEATH: CHRONIC IMMUNE ACTIVATION IN HIV PATHOGENESIS
- IS CHRONIC IMMUNE ACTIVATION ALONE ENOUGH TO INDUCE CD4+ T CELL LOSS?
- IMMUNE RECONSTITUTION INFLAMMATORY SYNDROME (IRIS)
- HOST IMMUNE CELL INFECTION WITH HIV
- Immunity and Immune-Based Diagnostic Tests
- 11 Immune Activation
- MAP Immune Responses in Crohn’s Disease Patients
- DETERMINANTS OF IMMUNE RESPONSE
- MARKERS OF IMMUNE ACTIVATION
- AUTOLOGOUS CELL-MEDIATED KILLING RESULTS IN IMMUNE SUPPRESSION