Net secondary production is equal to heterotroph growth
Not all of the organic matter consumed by heterotrophs is incorporated into heterotroph biomass. Some is used in respiration, and some is egested (lost in urine and feces). Net secondary production is the balance among ingestion, respiratory loss, and egestion:
net secondary production = ingestion — respiration — egestion
(20.2)
Net secondary production by a heterotroph depends on the quality of its food, related to its digestibility and nutrient content.
In addition, the physiology of the heterotroph influences how effectively its food intake is channeled into growth. Animals with high respiration rates (e.g., endotherms) have less energy left over to allocate to growth.Net secondary production in most terrestrial ecosystems is a small fraction of NPP, because of predation on herbivores, plant defenses, and the low nutrient content of many plants, as we'll see in Chapter 21. Net secondary production represents a greater fraction of NPP in aquatic ecosystems than it does in terrestrial ecosystems. The majority of net secondary production in most ecosystems is associated with detritivores, primarily bacteria and fungi.
A Case Study Revisited
Life in the Deep Blue Sea, How Can It Be?
In this chapter, we have emphasized the importance of photosynthetic autotrophs as the source of energy for ecosystems, since the vast majority of the energy that enters ecosystems is derived from visible solar radiation. Here and in Chapter 5, however, we have alluded to another source of energy for ecosystems: chemosynthesis. Some bacteria can use chemicals such as hydrogen sulfide (H2S and related chemical forms, HS- and S2-) as electron donors to take up carbon dioxide and convert it into carbohydrates:
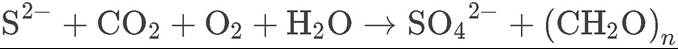
(20.3)
Bacteria that provide energy for ecosystems via chemosynthesis are known as chemoautotrophs.
The existence of Chemoautotrophic bacteria was known for at least a century before the discovery of hydrothermal vents, but their role in providing energy for the vent communities was uncertain.Initially, hypotheses suggested that the high velocity of water flow around the hydrothermal vents helped direct organic matter from the photic zone toward the filter-feeding invertebrates. However, several lines of evidence suggested that chemoautotrophs were the major source of energy for these ecosystems. First, the carbon isotopic ratios (13C∕12C) in the bodies of the vent invertebrates were different from those of phytoplankton in the photic zone (see Ecological Toolkit 5.1). Second, the tube worms collected from the vents (Riftia spp.) lacked mouths and digestive systems. These gutless tube worms also had structures called trophosomes, made up of highly vascularized tissues with specialized cells containing large amounts of bacteria (FIGURE 20.19). Elemental sulfur was found in the trophosomes, suggesting that sulfides were being chemically transformed in the tube worms' bodies. Enzymes associated with the Calvin cycle, the biochemical pathway used by autotrophs to synthesize carbohydrates (see Concept 5.2), as well as enzymes involved in sulfur metabolism were found in the trophosomes. Furthermore, the clams and other mollusks collected from the vent communities lacked some of the critical tissues for filter feeding, and they also had large amounts of bacteria in specialized tissues, as well as enzymes associated with
the Calvin cycle.
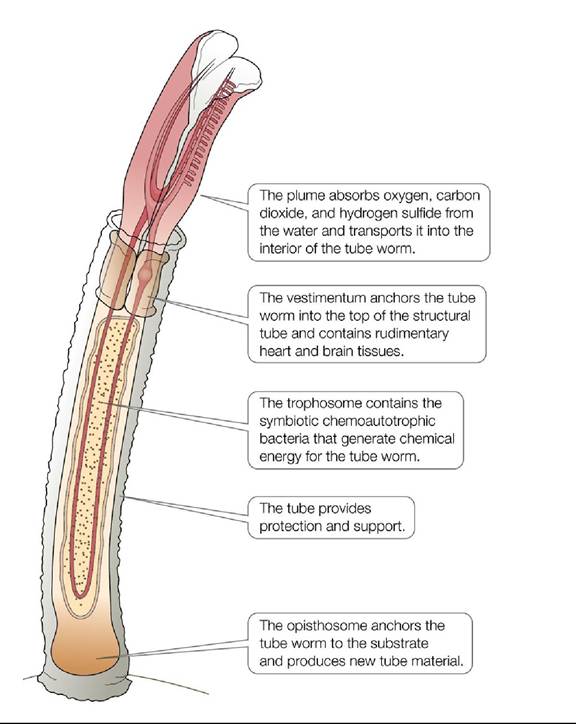
FIGURE 20.19 Riftia Anatomy Riftia tube worms have a number of specialized structures that make them well adapted to their hydrothermal vent environment. View larger image
All of this evidence pointed to the conclusion that deep-sea hydrothermal vent communities derive their energy from chemoautotrophic bacteria.
These bacteria also aid in detoxifying the sulfides in the water, which would normally inhibit aerobic respiration. Many of the abundant ocean vent organisms have symbiotic relationships with the bacteria—that is, they house the chemoautotrophs in their bodies, often in specialized structures. Is this interaction a mutualistic symbiosis of the kind described in Chapter 15? The tube worms and clams housing the bacteria benefit by obtaining carbohydrates to fuel their metabolic processes, growth, and reproduction, as well as from detoxification of the sulfides. Do the bacteria derive any benefit from the invertebrates? The answer is yes: the invertebrates provide them with a chemical environment unlike that found in the surrounding water, supplying them with more carbon dioxide, oxygen, and sulfides than they could obtain if they were free-living in the water or the sediments surrounding the vent. The symbiosis between the bacteria and the invertebrates is therefore mutualistic, resulting in higher productivity than if the organisms lived separately.Connections in Nature
Energy-Driven Succession and Evolution in Hydrothermal Vent Communities
Hydrothermal vent environments are dynamic, born with the eruption of new hot springs, which eventually cease to emit sulfide-laden water as the subsurface water channels are altered and the underlying magma cools (Van Dover 2000). When the hot springs no longer emit water, and the sulfide in the seawater has been consumed, the communities surrounding the vents collapse as their energy source disappears and the physical substrate falls apart. The life span of vent communities varies from approximately 20 to 200 years. Studies of colonization and development in these communities over the past 3 decades have provided insights into succession in marine communities in general (see Chapter 17 for a
general discussion of succession).
Succession in hydrothermal vent communities is relatively rapid and can be observed by periodically revisiting specific vents (FIGURE 20.20).
Although the logistic difficulty and expense of such investigations has limited the number of observations, some general trends have emerged. The rates of colonization and development of hydrothermal vent communities are higher when they are closer to other existing vent communities, as we might predict based on the theory of island biogeography (see Concept 18.3). Because the community’s energy is derived from chemosynthesis, colonization begins with chemoautotrophic bacteria, sometimes in numbers large enough to cloud the water. Tube worms are often the first invertebrates to arrive. Clams and other mollusks are thought to be stronger competitors for sites with optimal temperatures and water chemistry, and over time they increase in abundance at the expense of the tube worms. A few scavengers and carnivores, such as crabs and lobsters, are found in the developing community, although at low abundances. As the tube worm and bivalve populations decline with the drop in sulfide input when water flow from the vent decreases, the abundance of scavenger organisms increases until the energy available in the form of detritus is gone.
FIGURE 20.20 SuccessioninHydrothermalVentCommunities Species composition and abundances in a hydrothermal vent community change over time following the eruption of a hot spring. (From T. M. Shank et al. 1998. Deep-Sea Res II 45: 465-515.) View larger image
The pattern of succession in hydrothermal vent communities is subject to the same random factors that influence succession in other habitats: the order of arrival of organisms at a site can influence the long-term dynamics of the community (see Concept 17.4). Neighboring vent communities found in the same area of a mid-ocean ridge may show different stages of succession, associated with the stages of hot spring development, as well as different trajectories of succession due to differences in the organisms present.
Thus, collections of hydrothermal vents within the same general area are a mosaic of communities at different successional stages, similar to those in terrestrial forest patches, albeit separated by greater distances than the patches within a forest.The unique nature of the energy supply in hydrothermal vent communities would suggest strong evolutionary divergence between the organisms that inhabit the vents and their nearest non-vent relatives (see Concept 6.4). Where phylogenetic relationships between the vent organisms and their non-vent relatives have been worked out, the divergence is indeed deep, usually at the level of genus, family, or order. Since the discovery of hydrothermal vents, approximately 500 new vent species have been described; of these species, about 90% are endemic to hydrothermal vents. However, large areas of mid-ocean ridges potentially containing hydrothermal vents have yet to be explored.
The close association between the chemoautotrophic bacteria and their invertebrate hosts suggests the potential for a coevolutionary relationship of the type described in Concept 15.1. Have the invertebrates and their chemosynthetic bacterial symbionts evolved in concert? To address this question, Andrew Peek and colleagues compared the evolutionary relationships (phylogenetic trees; see Figure 6.16) of vent-dwelling clams in the family Vesicomyidae with those of their symbiotic bacteria (Peek et al. 1998). Clams in this family transfer bacteria to their offspring in the cytoplasm of their eggs. Peek and colleagues collected eight species of clams in three genera from hydrothermal vent communities at latitudes ranging from 18°N to 47°N and at depths ranging from 500 to 6,370 m. Ribosomal DNA taken from the clams and the bacteria was used to construct the phylogenetic trees. The two trees showed remarkable congruence (FIGURE 20.21),
providing strong evidence that speciation in the clams and in their bacterial symbionts has occurred synchronously.
Other vent groups lack this apparent coevolutionary relationship, however. For example, three different species of tube worms found in different geographic locations have been found to contain the same species of sulfur-oxidizing bacteria.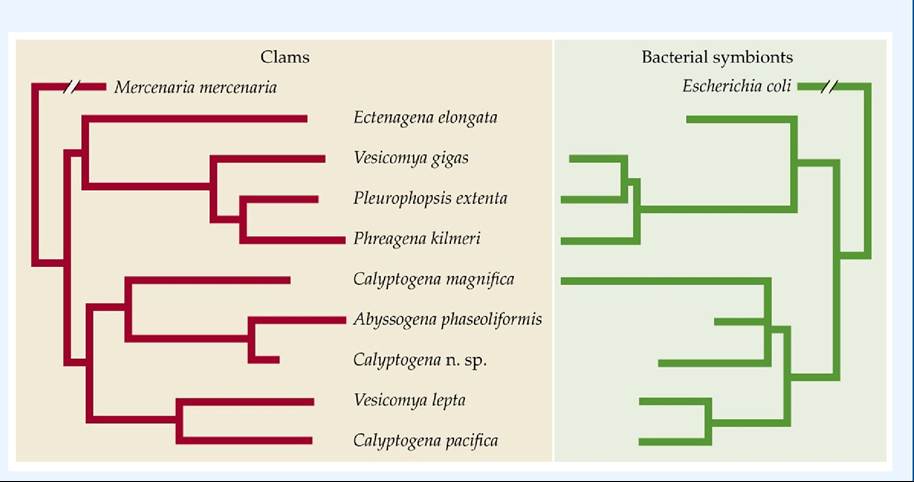
FIGURE 20.21 Coevolution of Vent Clams and Their Symbiotic Bacteria The phylogenetic trees of vesicomyid clams collected from hydrothermal vents and their accompanying chemoautotrophic bacterial symbionts show remarkable parallels, suggesting that these species have coevolved. (After A. S. Peek et al. 1998. Proc Natl Acad Sci USA 95: 9962-9966. © 1998 National Academy of Sciences, U.S.A.) View larger image
Some researchers have suggested that hydrothermal vents are a potential site for the origin of life on Earth. The reducing (i.e., electron-donating) geochemical environment of hydrothermal vents is conducive to the abiotic synthesis of amino acids, which would have been required for the development of living systems. Although amino acids are not stable in ocean water under the high pressures and temperatures found at some deep-sea hydrothermal vents, there are vents with lower temperatures at shallower depths where amino acid genesis could (and does) occur. As Cyndy Lee Van Dover (2000) so eloquently stated, “Vent water may be the ultimate soup in the sorcerer’s kettle.”
More on the topic Net secondary production is equal to heterotroph growth:
- Net secondary production is equal to heterotroph growth
- Energy that is derived from the consumption of organic compounds produced by other organisms is known as secondary production.
- NET EXPOSURE
- Net primary production is the energy remaining after respiratory losses
- Heterotroph diets can be determined from the isotopic composition of food sources
- SUMMARY
- New production
- Terrestrial and oceanic NPP are nearly equal
- Production of Simultaneous Rulership
- Pathogenesis of Secondary Otitis Media