ALL REVVED UP WITH NO PLACE TO GROW—ACTIVATION- INDUCED CELL DEATH (AICD) and peripheral deletion
Peripheral lymphocyte populations are regulated by homeostasis, deletion, and homeostatic proliferation. Peripheral T cells undergo rapid and extensive expansion upon activation.
In order to reduce lymphocyte numbers postinfection, T cells are subject to a number of selective pressures to reduce their numbers, leaving a small memory population. Some lymphocytes die following a suboptimal stimulation, either by weak MHC[Ag]:TCR interactions or lack of co-receptor involvement, and enter apoptosis during abortive proliferation. Some survive and constitute the memory pool of responders to subsequent challenge, but most activated lymphocytes undergo apoptosis in the postexpansion phase of the immune response.Mature peripheral T cells expand upon first encounter with antigen and undergo apoptosis upon secondary stimulation by a process known as activation-induced cell death (AICD).28 Resting lymphocytes are generally refractory to apoptotic stimuli, often because they do not express the necessary components to engage death pathways. Presumably, resting lymphocytes need not be sensitive to apoptotic stimuli, because they do not represent a significant risk, whereas activated lymphocytes, if misdirected, pose a substantial risk. The biochemical machinery necessary to drive apoptosis is expressed in peripheral T cells upon activation, but activation alone is not sufficient to drive a cell into apoptosis. AICD can be recapitulated in vitro by stimulating lymphocytes through the T cell receptor (TCR) in the presence of IL-2. Secondary stimulation 3 to 4 days later causes activated T cells to undergo AICD by a mechanism mediated, at least in part, by the c-myc-induced expression of Fas-ligand and engagement of the Fas-dependent extrinsic pathway. Induction of Fas- ligand is not sufficient to drive AICD; there is a corresponding sensitization of T cells to Fas- mediated apoptosis that occurs subsequent to the initial activation events.
Susceptibility to receptor- mediated apoptosis includes downregulation of the Fas inhibitory protein, c-FLIP (see Figure 5.3). Induction of Fas-ligand expression and subsequent ligand-receptor interactions have also been shown to occur in lymphocyte apoptosis independent of TCR engagement: genotoxic stress such as ultraviolet (UV) radiation, etoposide, and teniposide have been shown to cause an upregulation of Fas-ligand, which leads to lymphocyte apoptosis.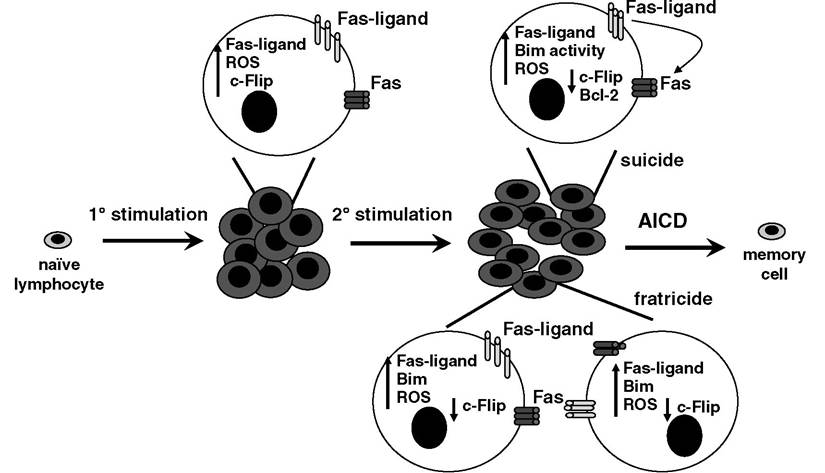
FIGURE 5.3 After a primary encounter with antigen, T lymphocytes expand; upregulate Fas, Fas-ligand, c- Flip, and Bim; and produce ROS. Upon secondary stimulation, a decrease of c-Flip levels is accompanied by an increase in sensitivity to apoptotic stimuli, notably Fas and Bim, key mediators of the extrinsic and intrinsic pathways, respectively. Cell autonomous suicide and fratricide-based mechanisms are employed to delete the majority of active lymphocytes, leaving a functional memory population. There are likely to be differences between CD4+ and CD8+ cells with respect to the contribution of each pathway, and there may also be subtleties in the employment of each mechanism, depending on the nature of the original antigenic stimulation.
Despite the demonstrated involvement of Fas-ligand and Fas in peripheral deletion and AICD, they are likely not the only mediators of apoptosis involved in this process. TNFα, a member of the same protein family of death ligands as Fas-ligand, has been implicated in both AICD and peripheral deletion.29,30 Activated lymphocytes are also sensitive to TRAIL-induced apoptosis, and the ligand is expressed upon activation. A definitive role for TRAIL in AICD has yet to be established.
As discussed above, the intrinsic apoptotic pathway can be activated in response to cellular stress, including growth factor withdrawal. Peripheral T cells are dependent on IL-2 for continued survival, and the BH3-only protein Bim is required to signal for apoptosis from IL-2 withdrawal in vitro.31 The ability of Bim to serve as a cellular sensor for IL-2 withdrawal may represent the downregulation of an immune response when growth factors become depleted due to usage or removal due to reduced production.
There are conflicting reports regarding the expression of Bim in activated T cells.
It was shown that Bim levels do not change in Vβ8+ cells after challenge with superantigen SEB (Staphylococcus enterotoxin B), but the activity of Bim was enhanced by a decrease in Bcl-2 protein levels.32 The net result was that Vβ8+ T cells were deleted in a Bim-dependent manner. A recent report showed differences in Bim levels in resting and activated CD4+ and CD8+ T cells33; CD4+ cells showed little change in Bim levels, whereas a more pronounced induction was observed in activated CD8+ cells. Therefore, it is possible that induction of Bim with or without a corresponding reduction of Bcl-2 may occur during peripheral deletion in a bulk population of SEB-responsive Vβ8+ cells.An interesting observation arose from early studies of Fas-ligand in AICD concerning the concept of cell autonomous apoptosis. Initial studies reported that a cell could undergo AICD in conditions devoid of other cells (i.e., cell autonomous death or “suicide”) (see Figure 5.3). Such a mechanism is necessary for negative selection when cells die in the absence of an appropriate signal.28,34 In some cases of autonomous AICD in vitro using T cell hybridomas, apoptosis was found to be dependent on Fas-Fas-ligand interactions. AICD need not be autonomous, as there are also examples using T cell hybridomas that require cell-cell contact to undergo AICD (i.e., “fratricide”).
Clearly, it would pose a problem if a lymphocyte were simply to die when activated. The coordinated expression of death proteins is tempered with the induction of death inhibitors. For example, lymphocytes express key components of the death machinery, including death ligands and receptors, notably, Fas-ligand/Fas and TRAIL/DR5, but they also induce expression of inhibitors of apoptosis, such as c-Flip, Bcl-2, and Bcl-xL. Why the apoptotic proteins are expressed early is not entirely clear, but there is some suggestion that proteins, such as FADD, and caspase-8 are required for activation and proliferation of T cells as well as for apoptosis.
Expression of Fas, FADD, and caspase-8 early during T cell activation may also represent a fail-safe mechanism that ensures that a cell is triggered to die if something goes awry. Presumably, it is better to remove an activated lymphocyte too soon than to have it persist as a rogue lymphocyte in the periphery, where it might cause significant harm.Resting lymphocytes are transcriptionally and metabolically inactive. Signaling by the T cell receptor (TCR) results in a complex kinase-dependent signal transduction cascade and activation of the T cell. The transcriptional machinery becomes increasingly active, and enhanced metabolic activity supports the growing needs of the activated lymphocyte for energy and growth. Reactive oxygen species (ROS), produced as a by-product during the heightened metabolic activity, were linked in both causal and consequential roles in programmed cell death. ROS have been implicated in AICD based on the observation that MnTBAP,35 an ROS scavenger, is capable of rescuing activated lymphocytes from death in culture ex vivo. Although this is a long way from proof that ROS is a causal factor in AICD, it has been since suggested that ROS produced by activated T cells are necessary for the upregulation of Fas-ligand, which was previously identified as a key factor in cell autonomous AICD.
More on the topic ALL REVVED UP WITH NO PLACE TO GROW—ACTIVATION- INDUCED CELL DEATH (AICD) and peripheral deletion:
- APPROPRIATE T CELL DEATH: CHRONIC IMMUNE ACTIVATION IN HIV PATHOGENESIS
- CLINICAL SIGNIFICANCE OF THE CELL DEATH INDUCED BY PR
- 17 Nonapoptotic HIV-Induced T Cell Death
- MODULATION OF HIV-MEDIATED IMMUNE ACTIVATION, AICD, AND EFFECT OF ANTIRETROVIRAL THERAPY
- MURDER BY PROXY—NONLYMPHOID CELLS IN PERIPHERAL LYMPHOCYTE DELETION
- CD4 AND CD8 T LYMPHOCYTES DURING HIV-INDUCED CHRONIC IMMUNE ACTIVATION
- IS CHRONIC IMMUNE ACTIVATION ALONE ENOUGH TO INDUCE CD4+ T CELL LOSS?
- OTHER FORMS OF CELL DEATH: NECROSIS AND AUTOPHAGY
- Section 3 Clinical Consequences of HIV- Induced Cell Deat
- CELL DEATH PATHWAY DURING ACUTE HIV-1 INFECTION
- INAPPROPRIATE T CELL APOPTOSIS: DIRECT HIV-INDUCED Cytopathicity and cd4 cross-linking
- HIV PR AND CELL DEATH
- CELL DEATH BY MURDER
- VPR MEDIATES CELL DEATH
- CELL DEATH BY SUICIDE
- VIRAL MEDIATORS OF DIRECT, NONAPOPTOTIC CELL DEATH