VPR MEDIATES CELL DEATH
Apoptotic vs. Necrotic Death
Maintenance of proper cell number requires a balance between proliferation, production, and programmed cell death (PCD). Inhibition of apoptosis leads to autoimmune disease and development of malignancies.
Conversely, enhanced apoptosis can cause diseases associated with neurodegeneration, ischemia-reperfusion damage, and immunodeficiency. The progressive decline in CD4+ T cells is the central feature of HIV infection and was proposed to be the result of increased levels of PCD (reviewed in Badley, Pilon, Landay, and Lynch72). The mechanism by which HIV infection causes lymphocyte destruction continues to be a matter of great debate. The earliest studies examining the mode of T cell death proposed that apoptosis was the predominant pathway involved. However, a recent study indicates that T cell destruction and the pathogenesis of HIV may involve an alternative form of necrotic cell death that does not rely on caspase activity.73,74 A large part of the disparate data stems from the highly divergent tissues, cells, models, and assays used to analyze PCD. For example, HIV replication was shown to lead to rapid death of peripherally infected T cells, thereby resulting in a constant turnover of lymphocytes.75,76 In contrast, analysis of lymph nodes showed that cell death occurs mainly in uninfected cells.77Almost all the HIV-1 proteins were implicated in modulating the apoptotic response of T cells. However, genetic analyses that directly correlate with the apoptosis-inducing activities of these proteins are lacking and raise significant concern as to the physiological involvement of these proteins as mediators of T cell death. Vpr is not a unique protein in this regard, although an emerging pool of literature has documented naturally occurring mutations in Vpr that directly abrogate cell death both in vitro and in vivo.
There are also conflicting reports that suggest that Vpr may serve as both an antiapoptotic and a proapoptotic regulator of T cell death. In this section, we will discuss the current models of apoptotic and nonapoptotic modes of Vpr-induced T cell death and new modes for inhibiting the pathophysiological effects of Vpr.Early studies on Vpr function initially found that expression of Vpr-induced myocytes to differentiate78 and led to reports demonstrating that the Vpr’s action was mediated by the glucocorticoid receptor complex.79,80 Given that glucocorticoids inhibit the action of NF-κB, Vpr was proposed to modify cellular functions by inducing a glucocorticoid effect, perhaps through modulation of T cell death or survival. Answers to how Vpr induced cell death came from studies transfecting Vpr into cells. In these cells, Vpr potently suppressed immune activation through upregulation of IκBα, which inhibited NF-κB transcriptional activity and resulted in the induction of cell death.81 These initial results led several investigators to examine details of Vpr-induced cell death. As noted previously, Vpr induces cell-cycle arrest at the G2 stage of the cell cycle and subsequently results in apoptosis of a number of human cells, including fibroblasts, T cell lines, and primary cells.9-11,82-85 These observations were universally validated in a number of different systems. For example, in one study, adenovirus was used to introduce Vpr into HeLa cells in the absence of other HIV gene products.10 Adenovirus infection with Vpr led to G2/M cell-cycle arrest within 24 h after infection, and staining with annexin V revealed significant apoptosis with increased activation of caspase-9. Moreover, mice expressing a transgene for Vpr on a CD4 promoter exhibited enhanced apoptotic cell death with a marked decrease in T cells of the lymphoid organs and peripheral blood.86
The major components of apoptosis initiated via the intrinsic mitochondrial pathway were largely clarified in recent years (reviewed in Green and Reed87 and Green and Kroemer88).
A more detailed review of this pathway can be found in Chapter 2. Execution of cell death through this route occurs by causing the translocation and insertion of Bax or Bak into the mitochondria membrane, loss in membrane potential, and release of cytochrome c into the cytosol. After being released, cytochrome c binds to form a complex consisting of procaspase-9 and apoptotic activating factor 1 (Apaf-1). The formation of this multimeric complex results in cleavage of procaspase-9 to an active caspase- 9. This pathway requires the continued presence of adenosine triphosphate(ATP) in order to carry out these energetic dependent reactions (reviewed in Green and Reed87 and Green and Kroemer88).Using synthetic Vpr peptides, it was demonstrated that a major target of Vpr is the ANT, a component of the inner mitochondrial permeability transition pore complex.70 Vpr binding to ANT causes loss of membrane potential, release of cytochrome c, and activation of caspase-3 and caspase- 9.9,10,70,85,89 The ability of Vpr to induce cell death seems to be dependent on the C-terminal H(S∕F)RIG motif, as specific mutations in this region abrogate Vpr’s ability to bind ANT and also reduce the capacity to induce cell death.11,70,89
Given the important role of the mitochondria in the production of ATP, the nucleotide pool within the cell provides an indirect measure of cellular bioenergetics. NMR analysis of α, β, and γ31P in HeLa cells infected with adenovirus containing Vpr demonstrated a lower density of α31P than that of β31P. When this analysis was used to compare β31P in necrotic cells vs. adenovirus Vpr-infected cells, necrotic cells had no detectable β3lP, whereas adenoviral-infected cells contained significant levels (reviewed in Muthumani et al.90). Because the apoptotic cascade is an energydependent process and necrosis is associated with energy collapse, this result is consistent with Vpr-infected cells maintaining active metabolism and cytoplasmic integrity and providing further evidence that apoptosis is the main pathway in which Vpr executes cell death.
The mechanism of HIV-l induced death was called into question based on studies demonstrating that infection of Jurkat and H9 T cell lines failed to exhibit characteristics of classical apoptosis.73 This included assays that measured caspase activity and activation, DNA fragmentation, annexin V exposure, and loss of mitochondrial membrane potential. Cells overexpressing the antiapoptotic proteins Bcl-xL or v-FLIP failed to prevent death. Furthermore, no difference in cytopathogenicity to HIV-1 infection was observed in Jurkat cells deficient for RIP, caspase-8, and FADD, or when caspase inhibitors were used. Therefore, the authors concluded that a caspase and a mitochondrial-independent mechanism were responsible for the observed cell death. In a concurrent article, the same authors used electron microscopy to distinguish apoptotic vs. necrotic cell death based on cell morphology. In this case, VSV-G pseudotype viruses were engineered to contain various HIV-1 gene products and were then used to infected T cell lines. In line with the notion that nonapoptotic pathways are induced by HIV-1, the authors found that Env was not required for HIV-1-induced T cell death.74 The C-terminal of Vpr was found to induce cell death even in the absence of caspase-9, AIF, or Apaf-1.83 Also the overexpression of Bcl-2 does not protect against Vpr-induced death, lending support to the notion that T cell death may occur through multiple pathways.
There were also several reports that Vpr may function as an antiapoptotic protein. HepG2 cells undergo apoptosis after treatment with sorbitol, but cells expressing Vpr were largely protected against these effects.91 In another study, low levels of Vpr stably expressed in Jurkat cells were shown to have an impaired capacity to undergo apoptosis by cycloheximide, TNF-α, anti-Fas, or serum starvation. Furthermore, cells transfected with antisense Vpr oligonucleotides were more sensitive to these apoptotic stimuli.92 In a later study, the same group found that the protective effect of Vpr could be recapitulated even when cells were infected in vitro with HIV-1, although this was observed only during early time points after infection.93 At later time points, these infected cells ultimately underwent apoptosis.
Further evidence suggesting that Vpr prevents cell death comes from a study examining the effects of Jurkat cells expressing Vpr on cell adhesion properties.94 Both cell-cell and cell-matrix interactions are important for regulating locomotion, differentiation, homing, growth, and cell death. However, lack of matrix attachment can induce apoptosis through a process termed “anoikis.” Along these lines, constitutive expression of Vpr in Jurkat cells exhibits features distinct from mock infected cells. In particular, there was a dramatic increase in polymerized F-actin and improved rearrangement of filamentous stress fibers. In addition, surface expression of α5 and α6 integrins as well as cadherins was significantly increased compared with controls. Stimulation of cells with Fas or cycloheximide∕TNFα triggered a lower degree of apoptosis in Vpr-expression cells. Therefore, enhanced cell-matrix, adhesion, and spreading of Jurkat cells by Vpr favor increased integrin signaling associated with inhibition of apoptosis and cell survival.94Based on this evidence, Vpr may have a dual role in modulating the apoptotic response that is dependent on several factors, including the time of infection and the level of Vpr. It may be possible that low levels of Vpr early in infection results in protection from cell death that facilitates maximal viral replication, persistence, and viral survival. However, ultimately, at later time points, high levels of Vpr mediate the death of the host cell to support viral spreading and dissemination.
Vpr Polymorphisms
Given the multitude of functions attributed to Vpr, numerous studies have attempted to identify naturally occurring polymorphisms within Vpr that are associated with enhanced or attenuated Vpr function. Studies in macaques, chimpanzees, and human subjects underscore the important contribution of Vpr to disease progression. For example, macaques infected with Vpr-defective SIV strains do not exhibit progression to AIDS or have severely attenuated disease.95-97 In a human HIV-1 infected patient and chimpanzees infected with nonfunctional Vpr, slower disease progression was observed.98 Therefore, it is reasonable to suggest that disease progression could be attributed to Vpr function.
A unique group of patients collectively referred to as long-term nonprogressors (LTNPs) were identified as being HIV-1 infected, with high viral titers but low levels of CD4+ T cell death. Importantly, these patients did not progress to AIDS, although some reports suggested that several of these patients ultimately showed disease progression. A number of these patient cohorts exist and were studied extensively to isolate both host and viral genetic aberrations that may account for their favorable clinical outcome. Many Vpr mutations were documented that interfere with cell-cycle arrest, decreased phosphorylation, and nuclear localization (Figure 7.3). A major portion of Vpr’s cytotoxicity is found clustered within the C-terminal, consisting of a dodecapeptide domain (residues 71 to 82). Several studies have demonstrated that polymorphisms within this arginine-rich C-terminal are frequently observed in LTNPs.13,99,100 In these cases, such mutations abolish Vpr function on mitochondria-induced apoptosis, cell-cycle arrest, and nuclear localization, and they may explain decreased pathogenicity of viruses harboring these alleles. One study found a unique polymorphism, Q3R, in a LTNP. This mutation resulted in a premature stop codon that correlated with reduced ability to induce T cell apoptosis.101 A more direct role of Vpr in disease progression was demonstrated in a molecular analysis of an HIV-1-infected mother-child LTNP pair. Clustering of amino acid substitutions in the cytotoxic domain of Vpr correlated with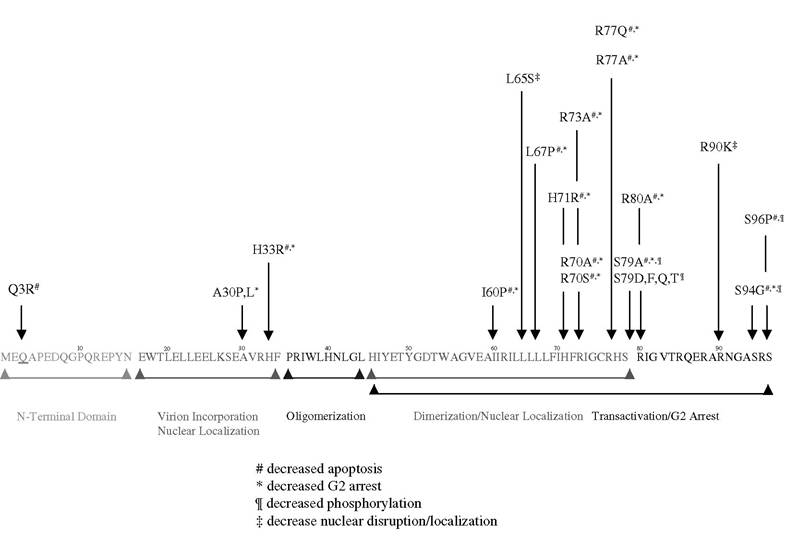
FIGURE 7.3 Mutations in Vpr domain affect its cellular activity.
high T cell count.13 However, reversion to wild-type Vpr resulted in a progressive decline in T cell numbers, strongly indicating that Vpr is an important determinant in viral pathogenesis and disease progression. In another study, arginine substitution at position 77 to glutamine was found at high frequency among LTNPs.89 This mutation was also observed in a larger sample of LTNP patients identified through the Los Alamos database and was associated with the reduced ability of Vpr to induce T cell death in vitro and in vivo. The incidence of this mutation was independently confirmed in another report.102 However, conflicting data were recently published as part of a larger clinical study examining the clinical outcomes of patients treated with a single- vs. a double-nucleoside reverse-transcriptase inhibitor.103 In this report, the authors found no correlation between the R77Q mutation and disease severity, progression, or absolute CD4+ T cell counts. Therefore, the role of Vpr mutations, specifically R77Q, was questioned as a predictor of clinical outcome. Several explanations may account for the contrasting observations. First, this study was designed to assess whether HAART masks any benefit toward disease progression that may have otherwise resulted from C-terminal mutations within the Vpr, whereas the original report studied only drug-naive HIV-1- infected patients with defined nondisease progression. Second, other viral or host factors that were suggested to play unique roles in disease progression were not defined in the latter study, whereas nef was found to be intact in all enrolled individuals, and no patient that was homozygous or heterozygous for the ∆32bp mutation in the chemokine co-receptor CCR5 was included in the former study. Third, viral load information was not available in the second study and may account for the lack of protective effects of the R77Q mutation. In addition, without viral load data, it would not be possible to speculate severity of pre-existing disease before treatment. Despite these discrepancies, 41% of the drug naive patients had viruses containing the R77Q mutation in Vpr, a value similar to that found for nearly 300 LTNPs in the Los Alamos database but lower than the 80% cited in the initial report. It is difficult to extract from the data in the second study whether patients who experienced an increase in CD4+ T cell counts were also drug naive. Perhaps the most compelling evidence will require the isolation of clinical viruses from both cohorts of patients so that careful in vitro analysis can be performed to assess the virulence of these mutant Vpr-containing isolates.
Novel Strategies to Inhibit Vpr Function
The multifunctional role of Vpr in the viral life cycle and its interaction with a number of host targets allow for the possibility of designing specific inhibitors to block Vpr function. Several line studies have demonstrated robust inhibition of Vpr activity in in vitro systems, and it will be important to validate these potential agents in animal models.
Cyclophilin blockers such as cyclosporine A and NIM811 were shown to have potent antiretroviral activity. For example, NIM811 interferes with nuclear localization of preintegration complexes. Cyclosporine was shown to block expression of interleukin-2, thereby exhibiting indirect antiviral activity through reducing T cell activation and release of infectious virions.104 In rhesus monkeys acutely infected with SIV, cyclosporine A showed some benefit in preventing T cell loss,105 but human trials in advanced stages of AIDS showed only a mild effect in patients treated with cyclosporine A. Because cyclophilin A inhibitors are less toxic, these drugs need to be evaluated clinically for and in combination with current HIV therapy.
Vpr-mediated cell-cycle arrest was observed in fission yeast systems and acts through a pathway involving PP2A, Wee1, and cdc25c.29,106 In addition, Vpr experimental evidence demonstrates that Vpr induces cell killing of budding yeast through its effect on the mitochondrial transition pore complex.70 Therefore, an elegant genetic system using the budding yeast S. cerevisiae was employed to screen for GST-fusion hexamic peptides, which suppressed the growth arrest phenotype of HIV-1 Vpr.107 In this system, yeast expression plasmids were generated, encoding Vpr under the control of galactose-inducible GAL1 promoter. When grown in Vpr-inducible medium, cells transformed with wild-type Vpr underwent growth arrest compared with control. When a hexamic peptide library consisting of 1 ? 108 GST peptides was transformed into yeast, 15 clones were found to exhibit strong growth defects. Sequence analysis revealed that all GST-inhibiting peptides contained a conserved diW motif (WXXF) and a stretch of hydrophobic residues at their C-terminals. This WXXF motif was previously shown to interact with Vpr,108 but the basis of this interaction is not known. Furthermore, these peptides interfered with the ability of VSV-G pseudotype viruses to induce G2 arrest and apoptosis. It was concluded that these diW-containing peptides may be capable of preventing protein-protein interactions between Vpr and host targets or may act as dissociative inhibitors. Along similar lines, other investigators examined the effects of dominant negative Vpr mutants on HIV-1 replication. Mutation of arginine residue 73 to serine in Vpr reduced the transactivation and G2 arrest by wild-type Vpr.109,110 However, the ability of this dominant negative R73S to block apoptosis was not investigated. Other approaches were to use antagonists of the glucocorticoid receptor (RU486) and pentoxifylline.79
Using another similar yeast genetic approach, one group screened for suppressors of G2 arrest (reviewed in Bukrinsky and Zhao111). Out of more than 2.0 ? 104 transformants, two small heatshock proteins, Hsp27 and Hsp70, were identified to inhibit G2 arrest by Vpr. When transfected into 293T cells, Hsp27 strongly suppressed G2 arrest induced by VSV-G pseudotype HIV-1 infection, whereas HSP70 had similar suppressive properties. The ability of HIV-1 to replicate in nonproliferating macrophages is dependent on the function of Vpr. It is believed that Vpr assists in the translocation of viral PICs via the nuclear envelope through interactions with a number of host proteins, including nucleoporins, importins, and heat-shock proteins. Hsp70 was found to stimulate nuclear import in a cell-free system.112 More recently, Hsp70 expression was induced following HIV-1 infection of macrophages.113 Interestingly, in the absence of Vpr, Hsp70 was able to stimulate import and replication in macrophages. However, Hsp70 reduced replication and reversed G2 arrest in Vpr- sufficient viruses. The authors concluded that Hsp70 and Vpr act in parallel pathways when expressed separately but inhibit each other’s activities when dually expressed. These results may provide an opportunity to modulate Hsp70 function during HIV-1 infection.
Recent work showed that the use of RNAi technology may have potential in inhibiting HIV-1 replication and other viral functions. Small interfering RNAs (siRNAs) are RNA duplexes of 21 to 23 nucleotides that induce specific degradation of a homologous mRNA that results in gene siliencing.114-116 One of the earliest findings that HIV-1 infection could be inhibited was the discovery that homozymous mutations in the chemokine co-receptors CCR5 conferred protection against HIV-1 infection.117 At least one other study showed that knockdown of CCR5 provided strong protection against infection in vitro,118 another potential target of RNAi during the viral uncoating and entry process of the target cell.
Another approach was to develop a system to deliver exogenous siRNAs through endogenous expression via DNA plasmids. This method would bypass the need to produce siRNA in cells, which have low expression of Dicer.119,120 Still, the requirement for stem-loop structure processing by Dicer or another endonuclease was problematic. Recent work showed that these stem-loop structures can be processed in a manner similar to micro-RNA precursors, and anti-HIV siRNAs processing from micro-RNA precursors can occur efficiently.121 Both HIV-1 Rev and Tat can be inhibited by these approaches in both transient transfection or from a lentiviral transduction of hematopoietic progenitor cells.122,123 In these studies, cotransfection of siRNA directed against Rev and Tat reduced the level of HIV-1 NL4-3 proviral DNA and inhibition of p24 expression by 4 logs123 in human T cell lines and primary lymphocytes.124 In similar work, silencing of CD4 blocked viral entry and syncytia formation.125 Given the proof of concept in these studies, it will be of interest to determine whether HIV-1 Vpr may be a potential target for siRNA silencing. Conceivably, the knockdown of Vpr expression would prevent infection of nondividing cells as well as abrogate its cell-cycle-inducing functions. Also, decreased Vpr expression may mimic naturally occurring mutations in Vpr, whereby these alleles have reduced capacity to induce cell death of target cells.
The advances in siRNA technology and continued improvement in its efficiency provide a unique opportunity to exploit delivery of anti-HIV treatments, including those that target Vpr. Loss- of-function Vpr studies indicate that inhibition of Vpr activity on cell cycle, apoptosis, or nuclear integration may be an alternative strategy to curb the deleterious effects of HIV-1 infection in humans.
REFERENCES
1. Cohen, E.A., Dehni, G., Sodroski, J.G., and Haseltine, W.A., Human immunodeficiency virus Vpr product is a virion-associated regulatory protein, J. Virol., 64(6), 3097-3099, 1990.
2. Zhao, Y. and Elder, R.T., Yeast perspectives on HIV-1 VPR, Front Biosci., 5(1), D905-D916, 2000.
3. Morellet, N., Bouaziz, S., Petitjean, P., and Roques, B.P., NMR structure of the HIV-1 regulatory protein VPR, J. Mol. Biol., 327(1), 215-227, 2003.
4. Engler, A., Stangler, T., and Willbold, D., Structure of human immunodeficiency virus type 1 Vpr(34-51) peptide in micelle containing aqueous solution, Eur. J. Biochem., 269(13), 3264-3269, 2002.
5. Luo, Z., Butcher, D.J., Murali, R., Srinivasan, A., and Huang, Z., Structural studies of synthetic peptide fragments derived from the HIV-1 Vpr protein, Biochem. Biophys. Res. Commun., 244(3), 732-736, 1998.
6. Wecker, K., Morellet, N., Bouaziz, S., and Roques, B.P., NMR structure of the HIV-1 regulatory protein Vpr in H2O/trifluoroethanol. Comparison with the Vpr N-terminal (1-51) and C-terminal (52-96) domains, Eur. J. Biochem., 269(15), 3779-3788, 2002.
7. Zhou, Y., Lu, Y., and Ratner, L., Arginine residues in the C-terminus of HIV-1 Vpr are important for nuclear localization and cell cycle arrest, Virology, 242(2), 414-424, 1998.
8. Zhou, Y. and Ratner, L., Phosphorylation of human immunodeficiency virus type 1 Vpr regulates cell cycle arrest, J. Virol., 74(14), 6520-6527, 2000.
9. Muthumani, K., Hwang, D.S., Desai, B.M., Zhang, D., Dayes, N., Green, D.R., and Weiner, D.B., HIV-1 Vpr induces apoptosis through caspase 9 in T cells and peripheral blood mononuclear cells, J. Biol. Chem., 277(40), 37820-37831, 2002.
10. Muthumani, K., Zhang, D., Hwang, D.S., Kudchodkar, S., Dayes, N.S., Desai, B.M., Malik, A.S., Yang, J.S., Chattergoon, M.A., Maguire, H.C., Jr., and Weiner, D.B., Adenovirus encoding HIV-1 Vpr activates caspase 9 and induces apoptotic cell death in both p53 positive and negative human tumor cell lines, Oncogene, 21(30), 4613-4625, 2002.
11. Jacotot, E., Ravagnan, L., Loeffler, M., Ferri, K.F., Vieira, H.L., Zamzami, N., Costantini, P., Druil- lennec, S., Hoebeke, J., Briand, J.P., Irinopoulou, T., Daugas, E., Susin, S.A., Cointe, D., Xie, Z.H., Reed, J.C., Roques, B.P., and Kroemer, G., The HIV-1 viral protein R induces apoptosis via a direct effect on the mitochondrial permeability transition pore, J. Exp. Med., 191(1), 33-46, 2000.
12. Wecker, K. and Roques, B.P., NMR structure of the (1-51) N-terminal domain of the HIV-1 regulatory protein Vpr, Eur. J. Biochem., 266(2), 359-369, 1999.
13. Wang, B., Ge, Y.C., Palasanthiran, P., Xiang, S.H., Ziegler, J., Dwyer, D.E., Randle, C., Dowton, D., Cunningham, A., and Saksena, N.K., Gene defects clustered at the C-terminus of the Vpr gene of HIV-1 in long-term nonprogressing mother and child pair: in vivo evolution of Vpr quasispecies in blood and plasma, Virology, 223(1), 224-232, 1996.
14. Tungaturthi, P.K., Sawaya, B.E., Ayyavoo, V., Murali, R., and Srinivasan, A., HIV-1 Vpr: genetic diversity and functional features from the perspective of structure, DNA Cell Biol., 23(4), 207-222, 2004.
15. Bukrinsky, M. and Adzhubei, A., Viral protein R of HIV-1, Rev. Med. Virol., 9(1), 39-49, 1999.
16. Singh, S.P., Tomkowicz, B., Lai, D., Cartas, M., Mahalingam, S., Kalyanaraman, V.S., Murali,
R., and Srinivasan, A., Functional role of residues corresponding to helical domain II (amino acids 35 to 46) of human immunodeficiency virus type 1 Vpr, J. Virol., 74(22), 10650-10657, 2000.
17. Mahalingam, S., Khan, S.A., Murali, R., Jabbar, M.A., Monken, C.E., Collman, R.G., and Srinivasan, A., Mutagenesis of the putative -helical domain of the Vpr protein of human immunodeficiency virus type 1: effect on stability and virion incorporation, Proc. Natl. Acad. Sci. U.S.A., 92(9), 3794-3798, 1995.
18. Mahalingam, S., Khan, S.A., Jabbar, M.A., Monken, C.E., Collman, R.G., and Srinivasan, A., Identification of residues in the N-terminal acidic domain of HIV-1 Vpr essential for virion incorporation, Virology, 207(1), 297-302, 1995.
19. Mahalingam, S., Ayyavoo, V., Patel, M., Kieber-Emmons, T., and Weiner, D.B., Nuclear import, virion incorporation, and cell cycle arrest/differentiation are mediated by distinct functional domains of human immunodeficiency virus type 1 Vpr, J. Virol., 71(9), 6339-6347, 1997.
20. Chen, M., Elder, R.T., Yu, M., O’Gorman, M.G., Selig, L., Benarous, R., Yamamoto, A., and Zhao, Y., Mutational analysis of Vpr-induced G2 arrest, nuclear localization, and cell death in fission yeast, J. Virol., 73(4), 3236-3245, 1999.
21. Vodicka, M.A., Koepp, D.M., Silver, P.A., and Emerman, M., HIV-1 Vpr interacts with the nuclear transport pathway to promote macrophage infection, Genes Dev., 12(2), 175-185, 1998.
22. Nishizawa, M., Kamata, M., Katsumata, R., and Aida, Y., A carboxy-terminally truncated form of the human immunodeficiency virus type 1 Vpr protein induces apoptosis via G(1) cell cycle arrest, J. Virol., 74(13), 6058-6067, 2000.
23. Macreadie, I.G., Arunagiri, C.K., Hewish, D.R., White, J.F., and Azad, A.A., Extracellular addition of a domain of HIV-1 Vpr containing the amino acid sequence motif H(SZF)RIG causes cell membrane permeabilization and death, Mol. Microbiol., 19(6), 1185-1192, 1996.
24. Macreadie, I.G., Castelli, L.A., Hewish, D.R., Kirkpatrick, A., Ward, A.C., and Azad, A.A., A domain of human immunodeficiency virus type 1 Vpr containing repeated H(SZF)RIG amino acid motifs causes cell growth arrest and structural defects, Proc. Natl. Acad. Sci. U.S.A., 92(7), 2770-2774, 1995.
25. Agostini, I., Popov, S., Hao, T., Li, J.H., Dubrovsky, L., Chaika, O., Chaika, N., Lewis, R., and Bukrinsky, M., Phosphorylation of Vpr regulates HIV type 1 nuclear import and macrophage infection, AIDS Res. Hum. Retroviruses, 18(4), 283-288, 2002.
26. Lopez-Girona, A., Furnari, B., Mondesert, O., and Russell, P., Nuclear localization of Cdc25 is regulated by DNA damage and a 14-3-3 protein, Nature, 397(6715), 172-175, 1999.
27. Krek, W. and Nigg, E.A., Differential phosphorylation of vertebrate p34cdc2 kinase at the G1ZS and G2ZM transitions of the cell cycle: identification of major phosphorylation sites, EMBO J., 10(2), 305-316, 1991.
28. Zhao, Y., Cao, J., O’Gorman, M.R., Yu, M., and Yogev, R., Effect of human immunodeficiency virus type 1 protein R (Vpr) gene expression on basic cellular function of fission yeast Schizosaccharomyces pombe, J. Virol., 70(9), 5821-5826, 1996.
29. Elder, R.T., Yu, M., Chen, M., Edelson, S., and Zhao, Y., Cell cycle G2 arrest induced by HIV-1 Vpr in fission yeast (Schizosaccharomyces pombe) is independent of cell death and early genes in the DNA damage checkpoint, Virus Res., 68(2), 161-173, 2000.
30. He, J., Choe, S., Walker, R., Di Marzio, P., Morgan, D.O., and Landau, N.R., Human immunodeficiency virus type 1 viral protein R (Vpr) arrests cells in the G2 phase of the cell cycle by inhibiting p34cdc2 activity, J. Virol., 69(11), 6705-6711, 1995.
31. Furnari, B., Rhind, N., and Russell, P., Cdc25 mitotic inducer targeted by Chk1 DNA damage checkpoint kinase, Science, 277(5331), 1495-1497, 1997.
32. Masuda, M., Nagai, Y., Oshima, N., Tanaka, K., Murakami, H., Igarashi, H., and Okayama, H., Genetic studies with the fission yeast Schizosaccharomyces pombe suggest involvement of Wee1, Ppa2, and Rad24 in induction of cell cycle arrest by human immunodeficiency virus type 1 Vpr, J. Virol., 74(6), 2636-2646, 2000.
33. Heinzinger, N.K., Bukinsky, M.I., Haggerty, S.A., Ragland, A.M., Kewalramani, V., Lee, M.A., Gendelman, H.E., Ratner, L., Stevenson, M., and Emerman, M., The Vpr protein of human immunodeficiency virus type 1 influences nuclear localization of viral nucleic acids in nondividing host cells, Proc. Natl. Acad. Sci. U.S.A., 91(15), 7311-7315, 1994.
34. Popov, S., Rexach, M., Ratner, L., Blobel, G., and Bukrinsky, M., Viral protein R regulates docking of the HIV-1 preintegration complex to the nuclear pore complex, J. Biol. Chem., 273(21), 13347-13352, 1998.
35. Popov, S., Rexach, M., Zybarth, G., Reiling, N., Lee, M.A., Ratner, L., Lane, C.M., Moore, M.S., Blobel, G., and Bukrinsky, M., Viral protein R regulates nuclear import of the HIV-1 pre-integration complex, EMBO J., 17(4), 909-917, 1998.
36. Subbramanian, R.A., Kessous-Elbaz, A., Lodge, R., Forget, J., Yao, X.J., Bergeron, D., and Cohen, E.A., Human immunodeficiency virus type 1 Vpr is a positive regulator of viral transcription and infectivity in primary human macrophages, J. Exp. Med., 187(7), 1103-1111, 1998.
37. Emerman, M., HIV-1, Vpr and the cell cycle, Curr. Biol., 6(9), 1096-1103, 1996.
38. de Noronha, C.M., Sherman, M.P., Lin, H.W., Cavrois, M.V., Moir, R.D., Goldman, R.D., and Greene, W.C., Dynamic disruptions in nuclear envelope architecture and integrity induced by HIV-1 Vpr, Science, 294(5544), 1105-1108, 2001.
39. Gallay, P., Stitt, V., Mundy, C., Oettinger, M., and Trono, D., Role of the karyopherin pathway in human immunodeficiency virus type 1 nuclear import, J. Virol., 70(2), 1027-1032, 1996.
40. Gallay, P., Hope, T., Chin, D., and Trono, D., HIV-1 infection of nondividing cells through the recognition of integrase by the importin/karyopherin pathway, Proc. Natl. Acad. Sci. U.S.A., 94(18), 9825-9830, 1997.
41. Moore, J.D., Yang, J., Truant, R., and Kornbluth, S., Nuclear import of Cdk/cyclin complexes: identification of distinct mechanisms for import of Cdk2/cyclin E and Cdc2/cyclin B1, J. Cell Biol., 144(2), 213-224, 1999.
42. Lu, Y.L., Spearman, P., and Ratner, L., Human immunodeficiency virus type 1 viral protein R localization in infected cells and virions, J. Virol., 67(11), 6542-6550, 1993.
43. Sherman, M.P., de Noronha, C.M., Eckstein, L.A., Hataye, J., Mundt, P., Williams, S.A., Neidleman, J.A., Goldsmith, M.A., and Greene, W.C., Nuclear export of Vpr is required for efficient replication of human immunodeficiency virus type 1 in tissue macrophages, J. Virol., 77(13), 7582-7589, 2003.
44. Eckstein, D.A., Sherman, M.P., Penn, M.L., Chin, P.S., De Noronha, C.M., Greene, W.C., and Goldsmith, M.A., HIV-1 Vpr enhances viral burden by facilitating infection of tissue macrophages but not nondividing CD4+ T cells, J. Exp. Med., 194(10), 1407-1419, 2001.
45. Amini, S., Khalili, K., and Sawaya, B.E., Effect of HIV-1 Vpr on cell cycle regulators, DNA Cell Biol., 23(4), 249-260, 2004.
46. Jowett, J.B., Planelles, V., Poon, B., Shah, N.P., Chen, M.L., and Chen, I.S., The human immunodeficiency virus type 1 Vpr gene arrests infected T cells in the G2 + M phase of the cell cycle, J. Virol., 69(10), 6304-6313, 1995.
47. Moir, R.D., Spann, T.P., Herrmann, H., and Goldman, R.D., Disruption of nuclear lamin organization blocks the elongation phase of DNA replication, J. Cell Biol., 149(6), 1179-1192, 2000.
48. Cohen, J., New role for HIV: a vehicle for moving genes into cells, Science, 272(5259), 195. 1996.
49. Wang, L., Mukherjee, S., Jia, F., Narayan, O., and Zhao, L.J., Interaction of virion protein Vpr of human immunodeficiency virus type 1 with cellular transcription factor Sp1 and trans-activation of viral long terminal repeat, J. Biol. Chem., 270(43), 25564-25569, 1995.
50. Agostini, I., Navarro, J.M., Rey, F., Bouhamdan, M., Spire, B., Vigne, R., and Sire, J., The human immunodeficiency virus type 1 Vpr transactivator: cooperation with promoter-bound activator domains and binding to TFIIB, J. Mol. Biol., 261(5), 599-606, 1996.
51. Kino, T., Gragerov, A., Slobodskaya, O., Tsopanomichalou, M., Chrousos, G.P., and Pavlakis, G.N., Human immunodeficiency virus type 1 (HIV-1) accessory protein Vpr induces transcription of the HIV-1 and glucocorticoid-responsive promoters by binding directly to p300/CBP coactivators, J. Virol., 761(19), 9724-9734, 2002.
52. Mahalingam, S., Ayyavoo, V., Patel, M., Kieber-Emmons, T., Kao, G.D., Muschel, R.J., and Weiner, D.B., HIV-1 Vpr interacts with a human 34-kDa mov34 homologue, a cellular factor linked to the G2/M phase transition of the mammalian cell cycle, Proc. Natl. Acad. Sci. U.S.A., 95(7), 3419-3424, 1998.
53. Withers-Ward, E.S., Jowett, J.B., Stewart, S.A., Xie, Y.M., Garfinkel, A., Shibagaki, Y., Chow, S.A., Shah, N., Hanaoka, F., Sawitz, D.G., Armstrong, R.W., Souza, L.M., and Chen, I.S., Human immunodeficiency virus type 1 Vpr interacts with HHR23A, a cellular protein implicated in nucleotide excision DNA repair, J. Virol., 71(12), 9732-9742, 1997.
54. Dieckmann, T., Withers-Ward, E.S., Jarosinski, M.A., Liu, C.F., Chen, I.S., and Feigon, J., Structure of a human DNA repair protein UBA domain that interacts with HIV-1 Vpr, Nat. Struct. Biol., 5(12), 1042-1047, 1998.
55. Gragerov, A., Kino, T., Ilyina-Gragerova, G., Chrousos, G.P., and Pavlakis, G.N., HHR23A, the human homologue of the yeast repair protein RAD23, interacts specifically with Vpr protein and prevents cell cycle arrest but not the transcriptional effects of Vpr, Virology, 245(2), 323-330, 1998.
56. Mansky, L.M., Preveral, S., Le Rouzic, E., Bernard, L.C., Selig, L., Depienne, C., Benarous, R., and Benichou, S., Interaction of human immunodeficiency virus type 1 Vpr with the HHR23A DNA repair protein does not correlate with multiple biological functions of Vpr, Virology, 282(1), 176-185, 2001.
57. Roux, P., Alfieri, C., Hrimech, M., Cohen, E.A., and Tanner, J.E., Activation of transcription factors NF-B and NF-IL-6 by human immunodeficiency virus type 1 protein R (Vpr) induces interleukin-8 expression, J. Virol., 74(10), 4658-4665, 2000.
58. Muthumani, K., Hwang, D.S., Dayes, N.S., Kim, J.J., and Weiner, D.B., The HIV-1 accessory gene Vpr can inhibit antigen-specific immune function, DNA Cell Biol., 21(9), 689-695, 2002.
59. Peng, C.Y., Graves, P.R., Thoma, R.S., Wu, Z., Shaw, A.S., and Piwnica-Worms, H., Mitotic and G2 checkpoint control: regulation of 14-3-3 protein binding by phosphorylation of Cdc25C on serine- 216, Science, 277(5331), 1501-1505, 1997.
60. Sanchez, Y., Wong, C., Thoma, R.S., Richman, R., Wu, Z., Piwnica-Worms, H., and Elledge, S.J., Conservation of the Chk1 checkpoint pathway in mammals: linkage of DNA damage to Cdk regulation through Cdc25, Science, 277(5331), 1497-1501, 1997.
61. Zeng, Y., Forbes, K.C., Wu, Z., Moreno, S., Piwnica-Worms, H., and Enoch, T., Replication checkpoint requires phosphorylation of the phosphatase Cdc25 by Cds1 or Chk1, Nature, 395(6701), 507-510, 1998.
62. Graves, P.R., Lovly, C.M., Uy, G.L., and Piwnica-Worms, H., Localization of human Cdc25C is regulated both by nuclear export and 14-3-3 protein binding, Oncogene, 20(15), 1839-1851, 2001.
63. Chen, L., Liu, T.H., and Walworth, N.C., Association of Chk1 with 14-3-3 proteins is stimulated by DNA damage, Genes Dev., 13(6), 675-685, 1999.
64. Chan, T.A., Hermeking, H., Lengauer, C., Kinzler, K.W., and Vogelstein, B., 14-3-3 is required to prevent mitotic catastrophe after DNA damage, Nature, 401(6753), 616-620, 1999.
65. Zander, K., Sherman, M.P., Tessmer, U., Bruns, K., Wray, V., Prechtel, A.T., Schubert, E., Henklein, P., Luban, J., Neidleman, J., Greene, W.C., and Schubert, U., Cyclophilin A interacts with HIV-1 Vpr and is required for its functional expression, J. Biol. Chem., 278(44), 43202-43213, 2003.
66. Luban, J., Absconding with the chaperone: essential cyclophilin-Gag interaction in HIV-1 virions, Cell, 87(7), 1157-1159, 1996.
67. Bouhamdan, M., Benichou, S., Rey, F., Navarro, J.M., Agostini, I., Spire, B., Camonis, J., Slupphaug, G., Vigne, R., Benarous, R., and Sire, J., Human immunodeficiency virus type 1 Vpr protein binds to the uracil DNA glycosylase DNA repair enzyme, J. Virol., 70(2), 697-704, 1996.
68. Selig, L., Benichou, S., Rogel, M.E., Wu, L.I., Vodicka, M.A., Sire, J., Benarous, R., and Emerman, M., Uracil DNA glycosylase specifically interacts with Vpr of both human immunodeficiency virus type 1 and simian immunodeficiency virus of sooty mangabeys, but binding does not correlate with cell cycle arrest, J. Virol., 71(6), 4842-4846, 1997.
69. Stark, L.A. and Hay, R.T., Human immunodeficiency virus type 1 (HIV-1) viral protein R (Vpr) interacts with Lys-tRNA synthetase: implications for priming of HIV-1 reverse transcription, J. Virol., 72(4), 3037-3044, 1998.
70. Jacotot, E., Ferri, K.F., El Hamel, C., Brenner, C., Druillennec, S., Hoebeke, J., Rustin, P., Metivier, D., Lenoir, C., Geuskens, M., Vieira, H.L., Loeffler, M., Belzacq, A.S., Briand, J.P., Zamzami, N., Edelman, L., Xie, Z.H., Reed, J.C., Roques, B.P., and Kroemer, G., Control of mitochondrial membrane permeabilization by adenine nucleotide translocator interacting with HIV-1 viral protein rR and Bcl-2, J. Exp. Med., 193(4), 509-519, 2001.
71. Phenix, B.N., Lum, J.J., Nie, Z., Sanchez-Dardon, J., and Badley, A.D., Antiapoptotic mechanism of HIV protease inhibitors: preventing mitochondrial transmembrane potential loss, Blood, 98(4), 1078-1085, 2001.
72. Badley, A.D., Pilon, A.A., Landay, A., and Lynch, D.H., Mechanisms of HIV-associated lymphocyte apoptosis, Blood, 96(9), 2951-2964, 2000.
73. Lenardo, M.J., Angleman, S.B., Bounkeua, V., Dimas, J., Duvall, M.G., Graubard, M.B., Hornung, F., Selkirk, M.C., Speirs, C.K., Trageser, C., Orenstein, J.O., and Bolton, D.L., Cytopathic killing of peripheral blood CD4(+) T lymphocytes by human immunodeficiency virus type 1 appears necrotic rather than apoptotic and does not require Env, J. Virol., 76(10), 5082-5093, 2002.
74. Bolton, D.L., Hahn, B.I., Park, E.A., Lehnhoff, L.L., Hornung, F., and Lenardo, M.J., Death of CD4(+) T-cell lines caused by human immunodeficiency virus type 1 does not depend on caspases or apoptosis, J. Virol., 76(10), 5094-5107, 2002.
75. Wei, X., Ghosh, S.K., Taylor, M.E., Johnson, V.A., Emini, E.A., Deutsch, P., Lifson, J.D., Bonhoeffer,
S., Nowak, M.A., Hahn, B.H., Saag, M.S., and Shaw, G.M., Viral dynamics in human immunodeficiency virus type 1 infection, Nature, 373(6510), 117-122, 1995.
76. Ho, D.D., HIV-1 dynamics in vivo, J. Biol. Regul. Homeost. Agents, 9(3), 76-77, 1995.
77. Finkel, T.H., Tudor-Williams, G., Banda, N.K., Cotton, M.F., Curiel, T., Monks, C., Baba, T.W., Ruprecht, R.M., and Kupfer, A., Apoptosis occurs predominantly in bystander cells and not in productively infected cells of HIV- and SIV-infected lymph nodes, Nat. Med., 1(2), 129-134, 1995.
78. Levy, D.N., Fernandes, L.S., Williams, W.V., and Weiner, D.B., Induction of cell differentiation by human immunodeficiency virus 1 Vpr, Cell, 72(4), 541-550, 1993.
79. Kino, T., Gragerov, A., Kopp, J.B., Stauber, R.H., Pavlakis, G.N., and Chrousos, G.P., The HIV-1 virion-associated protein Vpr is a coactivator of the human glucocorticoid receptor, J. Exp. Med., 189(1), 51-62, 1999.
80. Refaeli, Y., Levy, D.N., and Weiner, D.B., The glucocorticoid receptor type II complex is a target of the HIV-1 Vpr gene product, Proc. Natl. Acad. Sci. U.S.A., 92(8), 3621-3625, 1995.
81. Ayyavoo, V., Mahboubi, A., Mahalingam, S., Ramalingam, R., Kudchodkar, S., Williams, W.V., Green, D.R., and Weiner, D.B., HIV-1 Vpr suppresses immune activation and apoptosis through regulation of nuclear factor B [see comments], Nat. Med., 3(10), 1117-1123, 1997.
82. Levy, D.N., Refaeli, Y., MacGregor, R.R., and Weiner, D.B., Serum Vpr regulates productive infection and latency of human immunodeficiency virus type 1, Proc. Natl. Acad. Sci. U.S.A., 91(23), 10873-10877, 1994.
83. Roumier, T., Vieira, H.L., Castedo, M., Ferri, K.F., Boya, P., Andreau, K., Druillennec, S., Joza, N., Penninger, J.M., Roques, B., and Kroemer, G., The C-terminal moiety of HIV-1 Vpr induces cell death via a caspase-independent mitochondrial pathway, Cell Death Differ., 9(11), 1212-1219, 2002.
84. Stewart, S.A., Poon, B., Jowett, J.B., and Chen, I.S., Human immunodeficiency virus type 1 Vpr induces apoptosis following cell cycle arrest, J. Virol., 71(7), 5579-5592, 1997.
85. Stewart, S.A., Poon, B., Song, J.Y., and Chen, I.S., Human immunodeficiency virus type 1 Vpr induces apoptosis through caspase activation, J. Virol., 74(7), 3105-3111, 2000.
86. Yasuda, J., Miyao, T., Kamata, M., Aida, Y., and Iwakura, Y., T cell apoptosis causes peripheral T cell depletion in mice transgenic for the HIV-1 Vpr gene, Virology, 285(2), 181-192, 2001.
87. Green, D.R. and Reed, J.C., Mitochondria and apoptosis, Science, 281(5381), 1309-1312, 1998.
88. Green, D. and Kroemer, G., The central executioners of apoptosis: caspases or mitochondria?, Trends Cell Biol., 8(7), 267-271, 1998.
89. Lum, J.J., Cohen, O.J., Nie, Z., Weaver, J.G., Gomez, T.S., Yao, X.J., Lynch, D., Pilon, A.A., Hawley, N., Kim, J.E., Chen, Z., Montpetit, M., Sanchez-Dardon, J., Cohen, E.A., and Badley, A.D., Vpr R77Q is associated with long-term nonprogressive HIV infection and impaired induction of apoptosis, J. Clin. Invest., 111(10), 1547-1554, 2003.
90. Muthumani, K., Choo, A.Y., Hwang, D.S., Chattergoon, M.A., Dayes, N.N., Zhang, D., Lee, M.D., Duvvuri, U., and Weiner, D.B., Mechanism of HIV-1 viral protein R-induced apoptosis, Biochem. Biophys. Res. Commun., 304(3), 583-592, 2003.
91. Fukumori, T., Akari, H., Iida, S., Hata, S., Kagawa, S., Aida, Y., Koyama, A.H., and Adachi, A., The HIV-1 Vpr displays strong anti-apoptotic activity, FEBS Lett., 432(1-2), 17-20, 1998.
92. Conti, L., Rainaldi, G., Matarrese, P., Varano, B., Rivabene, R., Columba, S., Sato, A., Belardelli, F., Malorni, W., and Gessani, S., The HIV-1 Vpr protein acts as a negative regulator of apoptosis in a human lymphoblastoid T cell line: possible implications for the pathogenesis of AIDS, J. Exp. Med., 187(3), 403-413, 1998.
93. Conti, L., Matarrese, P., Varano, B., Gauzzi, M.C., Sato, A., Malorni, W., Belardelli, F., and Gessani, S., Dual role of the HIV-1 Vpr protein in the modulation of the apoptotic response of T cells, J. Immunol., 165(6), 3293-3300, 2000.
94. Matarrese, P., Conti, L., Varano, B., Gauzzi, M.C., Belardelli, F., Gessani, S., and Malorni, W., The HIV-1 Vpr protein induces anoikis-resistance by modulating cell adhesion process and microfilament system assembly, Cell Death Differ., 7(1), 25-36, 2000.
95. Gibbs, J.S., Lackner, A.A., Lang, S.M., Simon, M.A., Sehgal, P.K., Daniel, M.D., and Desrosiers, R.C., Progression to AIDS in the absence of a gene for Vpr or Vpx, J. Virol., 69(4), 2378-2383, 1995.
96. Hoch, J., Lang, S.M., Weeger, M., Stahl-Hennig, C., Coulibaly, C., Dittmer, U., Hunsmann, G., Fuchs, D., Muller, J., Sopper, S., Fleckenstein, B., and Ubuda, K.T., Vpr deletion mutant of simian immunodeficiency virus induces AIDS in rhesus monkeys, J. Virol. 69 (8), 4807-13, 1995.
97. Lang, S.M., Weeger, M., Stahl-Hennig, C., Coulibaly, C., Hunsmann, G., Muller, J., Muller-Her- melink, H., Fuchs, D., Wachter, H., Daniel, M.M., Duresiens, R.C., and Fleckenstein, B., Importance of Vpr for infection of rhesus monkeys with simian immunodeficiency virus, J. Virol., 67(2), 902-912, 1993.
98. Goh, W.C., Rogel, M.E., Kinsey, C.M., Michael, S.F., Fultz, P.N., Nowak, M.A., Hahn, B.H., and Emerman, M., HIV-1 Vpr increases viral expression by manipulation of the cell cycle: a mechanism for selection of Vpr in vivo, Nat. Med., 4(1), 65-71, 1998.
99. Saksena, N.K., Ge, Y.C., Wang, B., Xiang, S.H., Dwyer, D.E., Randle, C., Palasanthiran, P., Ziegler, J., and Cunningham, A.L., An HIV-1 infected long-term non-progressor (LTNP): molecular analysis of HIV-1 strains in the Vpr and Nef genes, Ann. Acad. Med. Singapore, 25(6), 848-854, 1996.
100. Yamada, T. and Iwamoto, A., Comparison of proviral accessory genes between long-term nonpro- gressors and progressors of human immunodeficiency virus type 1 infection, Arch. Virol., 145(5), 1021-1027, 2000.
101. Somasundaran, M., Sharkey, M., Brichacek, B., Luzuriaga, K., Emerman, M., Sullivan, J.L., and Stevenson, M., Evidence for a cytopathogenicity determinant in HIV-1 Vpr, Proc. Natl. Acad. Sci. U.S.A., 99(14), 9503-9508, 2002.
102. Rodes, B., Toro, C., Paxinos, E., Poveda, E., Martinez-Padial, M., Benito, J.M., Jimenez, V., Wrin,
T., Bassani, S., and Soriano, V., Differences in disease progression in a cohort of long-term non- progressors after more than 16 years of HIV-1 infection, AIDS, 18(8), 1109-1116, 2004.
103. Cavert, W., Webb, C.H., and Balfour, H.H., Jr., Alterations in the C-terminal region of the HIV-1 accessory gene Vpr do not confer clinical advantage to subjects receiving nucleoside antiretroviral therapy, J. Infect. Dis., 189(12), 2181-2184, 2004.
104. Emmel, E.A., Verweij, C.L., Durand, D.B., Higgins, K.M., Lacy, E., and Crabtree, G.R., Cyclosporin A specifically inhibits function of nuclear proteins involved in T cell activation, Science, 246(4937), 1617-1620, 1989.
105. Martin, L.N., Murphey-Corb, M., Mack, P., Baskin, G.B., Pantaleo, G., Vaccarezza, M., Fox, C.H., and Fauci, A.S., Cyclosporin A modulation of early virologic and immunologic events during primary simian immunodeficiency virus infection in rhesus monkeys, J. Infect. Dis., 176(2), 374-383, 1997.
106. Matsuda, Z., Yu, X., Yu, Q.C., Lee, T.H., and Essex, M., A virion-specific inhibitory molecule with therapeutic potential for human immunodeficiency virus type 1, Proc. Natl. Acad. Sci. U.S.A., 90(8), 3544-3548, 1993.
107. Yao, X.J., Lemay, J., Rougeau, N., Clement, M., Kurtz, S., Belhumeur, P., and Cohen, E.A., Genetic selection of peptide inhibitors of human immunodeficiency virus type 1 Vpr, J. Biol. Chem., 277(50), 48816-48826, 2002.
108. Gu, J., Emerman, M., and Sandmeyer, S., Small heat shock protein suppression of Vpr-induced cytoskeletal defects in budding yeast, Mol. Cell Biol., 17(7), 4033-4042, 1997.
109. Sawaya, B.E., Khalili, K., Rappaport, J., Serio, D., Chen, W., Srinivasan, A., and Amini, S., Suppression of HIV-1 transcription and replication by a Vpr mutant, Gene Ther., 6(5), 947-950, 1999.
110. Sawaya, B.E., Khalili, K., Gordon, J., Srinivasan, A., Richardson, M., Rappaport, J., and Amini, S., Transdominant activity of human immunodeficiency virus type 1 Vpr with a mutation at residue R73, J. Virol., 74(10), 4877-4881, 2000.
111. Bukrinsky, M. and Zhao, Y., Heat-shock proteins reverse the G2 arrest caused by HIV-1 viral protein R, DNA Cell Biol., 23(4), 223-225, 2004.
112. Agostini, I., Popov, S., Li, J., Dubrovsky, L., Hao, T., and Bukrinsky, M., Heat-shock protein 70 can replace viral protein R of HIV-1 during nuclear import of the viral preintegration complex, Exp. Cell Res., 259(2), 398-403, 2000.
113. Iordanskiy, S., Zhao, Y., DiMarzio, P., Agostini, I., Dubrovsky, L., and Bukrinsky, M., Heat-shock protein 70 exerts opposing effects on Vpr-dependent and Vpr-independent HIV-1 replication in macrophages, Blood, 104(6), 1367-1372, 2004.
114. Hutvagner, G. and Zamore, P.D., RNAi: nature abhors a double-strand, Curr. Opin. Genet. Dev., 12(2), 225-232, 2002.
115. Elbashir, S.M., Lendeckel, W., and Tuschl, T., RNA interference is mediated by 21- and 22-nucleotide RNAs, Genes Dev., 15(2), 188-200, 2001.
116. Elbashir, S.M., Harborth, J., Lendeckel, W., Yalcin, A., Weber, K., and Tuschl, T., Duplexes of 21- nucleotide RNAs mediate RNA interference in cultured mammalian cells, Nature, 411(6836), 494-498, 2001.
117. Doranz, F., Rucker, B.I., Liesnard, J., Farber, C., Saragosti, C.M., Lapoumeroulie, S., Samson, C., Liebert, M., Cognaux, J., Forceille, C., Muyldermans, G., Verhofstede, C., Burtonboy, G., Georges, M., Imai, T., Rana, S., Yi, Y., Smyth, R.J., Collman, R.G., Doms, R.W., Vassart, G., and Parmentier, M., Resistance to HIV-1 infection in caucasion individuals bearing mutant alleles of the CCR5 receptor gene, Nature, 382(6593), 722-725, 1996.
118. Martinez, M.A., Gutierrez, A., Armand-Ugon, M., Blanco, J., Parera, M., Gomez, J., Clotet, B., and Este, J.A., Suppression of chemokine receptor expression by RNA interference allows for inhibition of HIV-1 replication, Aids, 16(18), 2385-2390, 2002.
119. Kitabwalla, M. and Ruprecht, R.M., RNA interference—a new weapon against HIV and beyond, N. Engl. J. Med., 347(17), 1364-1367, 2002.
120. Jacque, J.M., Triques, K., and Stevenson, M., Modulation of HIV-1 replication by RNA interference, Nature, 418(6896), 435-438, 2002.
121. Zeng, Y. and Cullen, B.R., RNA interference in human cells is restricted to the cytoplasm, RNA, 8(7), 855-860, 2002.
122. Banerjea, A., Li, M.J., Bauer, G., Remling, L., Lee, N.S., Rossi, J., and Akkina, R., Inhibition of HIV- 1 by lentiviral vector-transduced siRNAs in T lymphocytes differentiated in SCID-hu mice and CD34+ progenitor cell-derived macrophages, Mol. Ther., 8(1), 62-71, 2003.
123. Lee, N.S., Dohjima, T., Bauer, G., Li, H., Li, M.J., Ehsani, A., Salvaterra, P., and Rossi, J., Expression of small interfering RNAs targeted against HIV-1 Rev transcripts in human cells, Nat. Biotechnol., 20(5), 500-505, 2002.
124. Coburn, G.A. and Cullen, B.R., Potent and specific inhibition of human immunodeficiency virus type 1 replication by RNA interference, J. Virol., 76(18), 9225-9231, 2002.
125. Novina, C.D., Murray, M.F., Dykxhoorn, D.M., Beresford, PJ., Riess, J., Lee, S.K., Collman, R.G., Lieberman, J., Shankar, P., and Sharp, P.A., siRNA-directed inhibition of HIV-1 infection, Nat. Med., 8(7), 681-686, 2002.
More on the topic VPR MEDIATES CELL DEATH:
- HIV KILLING OF INFECTED T CELLS
- Endocardial Spindle Cell Proliferation
- Badley A.D. (ed.). Cell Death During HIV Infection. Taylor & Francis,2006. — 511 p., 2006
- CELL DEATH PATHWAY DURING ACUTE HIV-1 INFECTION
- DEATH ASSOCIATED WITH GP120 BINDING TO CXCR4
- BIOLOGY OF VPR
- CONTENTS
- ALL REVVED UP WITH NO PLACE TO GROW—ACTIVATION- INDUCED CELL DEATH (AICD) and peripheral deletion
- ROLE OF APOPTOSIS IN T CELL DEVELOPMENT
- VIRAL MEDIATORS OF DIRECT, NONAPOPTOTIC CELL DEATH