Evidence of MAP in Animal- Derived Foods
The current evidence for MAP contamination of animal-derived foods, both raw and processed, is summarized in Table 2.1. The results presented are based on cultural, polymerase chain reaction (PCR or quantitative PCR) and phagebased detection of MAP.
The starting points for this summary table were the outcomes of a comprehensive review and meta-analysis of the evidence for MAP in animal-derived foods by Waddell et al. (2016); the additional information is from other published studies between 2011 and 2018. To the best of the author's knowledge, all published food surveillance studies have been included.2.2.1 Milk and dairy products
2.2.1.1 Raw milk
Raw cow's milk has been the focus of most MAP surveillance because it is recognized as a major factor in the transmission of paratuberculosis from cow to calf (Nielsen et al., 2008).
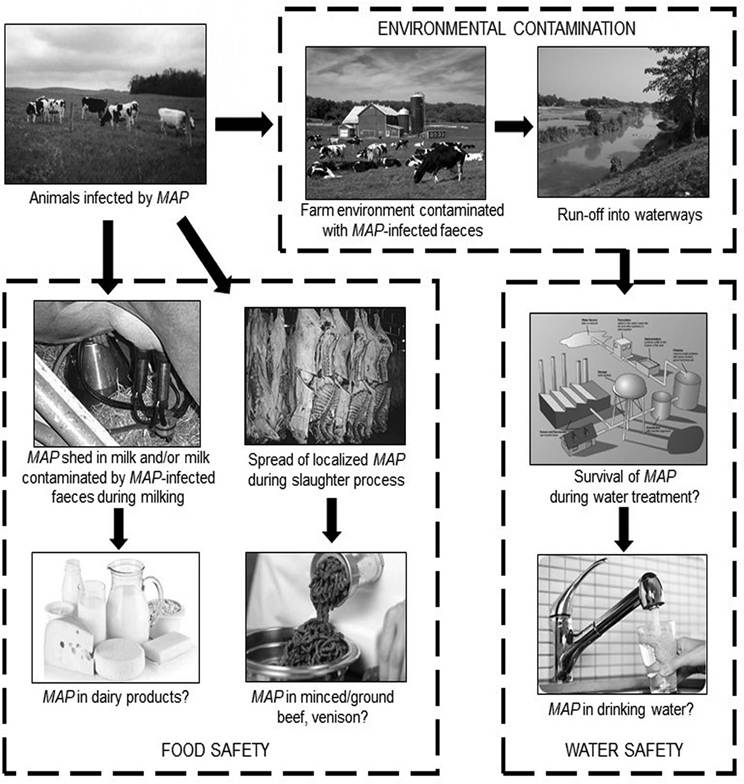
Fig. 2.1. Spread of Mycobacterium avium subsp. paratuberculosis (MAP) shed by infected animals in faeces and milk, and potential routes of human exposure to MAP via animal-derived foods and water.
Published studies have tested raw milk from individual animals, bulk tank at farm level or bulk silo milk prior to processing. Until the advent of quantitative real-time PCR and phage amplification assays for MAP, information on the levels of MAP in raw milk at these different points was limited. Early estimates of MAP levels in milk from sub-clinically affected cows derived by culture (e.g. Sweeney et al. (1992) reported two to eight colony-forming units (CFU) MAP/50 ml milk) are likely to have been underestimates for two reasons. First, chemical decontamination is generally applied to milk samples and this, depending on the method employed, negatively impacts the estimated counts of MAP (Dundee et al., 2001; Bradner et al., 2013).
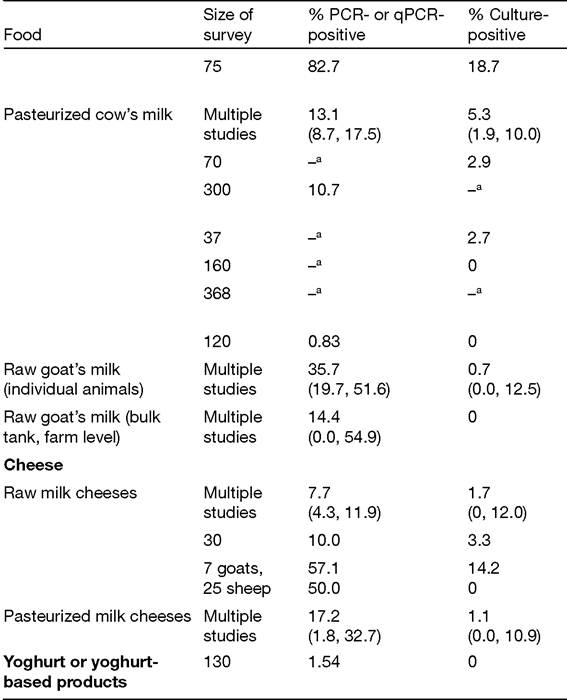
Second, in studies concerning milk from individual animals, the milk tested was obtained after thorough cleaning and disinfection of the exterior of the udder, removing the possibility of faecal contamination. Faecal contamination can and does occur during the milking process (Vissers et al., 2007), and different cleaning regimes applied to the udder affect the degree of faecal contaminationTable 2.1. A summary of the evidence for Mycobacterium avium subsp. paratuberculosis (MAP) contamination of raw and processed animal-derived foods obtained via surveillance studies. Findings of the meta-analysis by Waddell ef al. (2016) are listed first for each food category - milk, various dairy products and meat - followed by additional studies published since 2013.
5" class="lazyload" data-src="/files/uch_group75/uch_pgroup311/uch_uch7291/image/image005.jpg">
LR. Grant
Table 2.1. Continued
MAP in Animal-derived Foods and the Environment
anot tested or not reported.
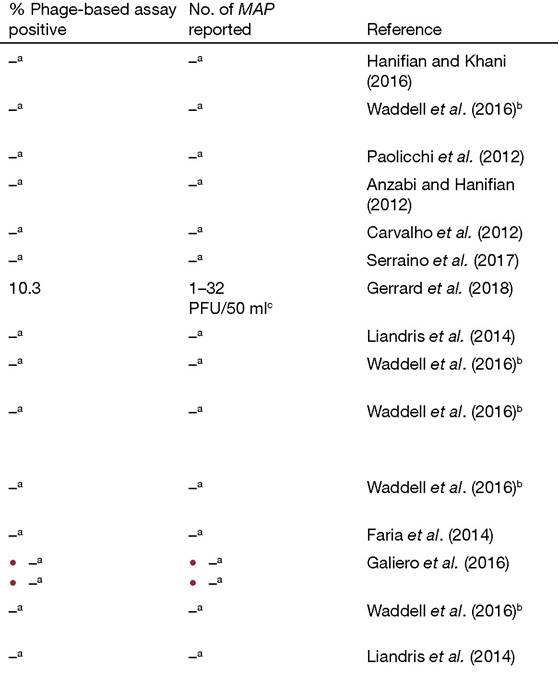
bFor data from Waddell etal. (2016), the figures are prevalence from a meta-analysis of multiple studies with 95% credible interval in brackets, ccounts determined by plaque assay; PFU is plaque-forming units.
dcounts determined by quantitative real-time polymerase chain reaction (qPCR).
I.R. Grant
(Gibson et al., 2008). It is clear from Table 2.1 that qPCR and phage-based detection of MAP in milk and dairy products consistently provide higher estimates of the proportion of samples positive for MAP than cultural methods; with percentage figures generally highest for phage assay results when all three methods are applied (Foddai and Grant, 2017; Grant et al., 2017).
While acknowledging that PCR does not differentiate between viable and dead bacteria, MAP cells detected in raw milk by qPCR are likely to be viable, and these numbers will not have been adversely affected by chemical decontamination (as CFU counts are). Phage amplification assays indicate numbers of only viable MAP cells present (by counting of plaque-forming units (PFU) produced), and, for this test also, the counts will not have been adversely affected by prior decontamination of the sample. The numbers of viable MAP in raw milk from individual animals and in bulk tank milk reported when qPCR and phagebased assays were employed (summarized in Table 2.1) indicate considerably higher levels in raw milk than previously thought.The level of MAP contamination of bulk tank milk at farm level is influenced by both infection status of the animals in a herd and hygiene practices employed during milking. Okura et al. (2013) used a modelling approach to estimate the concentration of MAP in bulk tank milk in dairy herds with within-herd Johne's disease prevalences of 7.5-60%; the estimated median loads of MAP were 0.54-7.53 CFU/ml (or 2 7-3 77CFU∕50ml). However, their model indicated that maximum concentration at a within-herd prevalence of 60% could be 1186 CFU/ml (or 59,300 CFU/50 ml) bulk tank milk, caused by shedding of high numbers of MAP in faeces of infected cows. Recently, Rani et al. (2019) used a Monte Carlo simulation model to predict the effect of hygiene practices (udder washing prior to milking, use of in-line milk filters, cleaning of milking equipment) on the levels of MAP contamination in bulk tank milk. Results indicated that 93% of MAP load in bulk tank milk is from faecal contamination, with the remainder contaminating via internal (direct shedding into milk) and environmental routes. Both Okura et al. (2013) and Rani et al. (2019) commented that a very few ‘super-shedding' animals in a herd may play a major role in raw milk MAP contamination levels.
The expectation has always been that there are two dilution steps for MAP in milk, towards very low or non-detectable levels, before processing: first, when milk from individual infected animals is mixed with milk of non-infected herdmates, and, second, when milk from one farm is mixed with milk from other farms before dairy processing. This may or may not be the case. Khol et al. (2013) reported the expected reduction in MAP levels from individual cow milks to bulk tank milk, whereas Hanifian and Khani (2016) reported a significant increase in MAP levels (p < 0.01) from ‘hygienically obtained' quarter milks to bulk tank. The latter authors concluded that MAP levels in raw milk are very dependent on the care and attention of farmers during the milking process (i.e. hygiene practices), in order to avoid faecal contamination.
Raw milk from sheep and goats has not been as extensively tested as cow's milk. In the developed world at least, these types of milk would tend to be consumed as dairy products rather than as liquid milk. The situation in developing countries is probably the reverse (Chaubey et al., 2017). There have been no recent surveillance studies of sheep and goat's milk. The review by Waddell et al. (2016) reported MAP mean PCR positivity of 35.7% in individual sheep and goat's milk and 14.4% in bulk tanks at farm level, but much lower culture positivity - 0.7% for individual milks only (Table 2.1). Greater difficulties in culturing isolates of MAP-S, as opposed to MAP-C (detailed in Chapters 12 and 18), may partly explain the low culture positivity reported for raw sheep or goat's milk surveys.
2.2.1.2 Pasteurized milk
The question of whether MAP can survive commercial pasteurization processes remains a contentious topic for the dairy industry (Robertson et al., 2017; Mullan, 2019). To date, 13 studies have reported the presence of viable MAP in retail pasteurized milk in different countries of the world. Waddell et al. (2016) included seven pasteurized milk surveys within their meta-analysis and reported mean prevalence of MAP of 13.1% by PCR (95% CI: 8.7, 17.5) and 5.3% by culture (95%CI: 1.9, 10.0).
Since then a further six pasteurized milk surveys have been reported in different parts of the world (Table 2.1), two of which reported isolation of viable MAP in 2.9% (Paolicchi etal., 2012) and 2.7% (Carvalho etal., 2012) of samples tested. A third study reported detection of viable MAP by the phage-PCR assay in 10.3% of retail semi-skimmed pasteurized milk samples in the UK (Gerrard et al., 2018), at levels of 1-32 PFU∕50ml milk in test positive samples. On the basis of the above surveillance data, it is difficult to conclude that humans are not being periodically exposed to some level of viable MAP via consumption of pasteurized milk in countries where paratuberculosis exists in dairy herds. However, it is still proving difficult for dairy processors to understand what might be the explanation for MAP survival when technical and engineering aspects of the milk pasteurization processes applied have been scrutinized and not found to be defective or deficient (Robertson et al., 2017; Mullan, 2019). The infective dose for humans is currently unknown, as is the potential risk that comes from repeated exposure to viable MAP.2.2.1.3 Various dairy products
Cheese (both raw and pasteurized milk varieties), powdered milk products (infant milk formula and calf milk replacer, specifically), and yoghurt or yoghurt-based products are the dairy products that have received attention as possible vehicles of human (or animal) exposure to MAP. Apart from a study of raw milk cheeses made from sheep's milk carried out in Italy (Galiero et al., 2016), MAP has consistently been isolated by culture (despite its recognized shortcomings) from raw and pasteurized milk cheeses, but PCR positivity of the cheeses was always much higher (Table 2.1). A single Greek study of 130 yoghurt or yoghurtbased products marketed for consumption by children (Liandris et al., 2014) found only PCR evidence of MAP presence (1.54%), but no MAP was cultured. There have been two reports of isolation of viable MAP in surveys of powdered milk products to date.
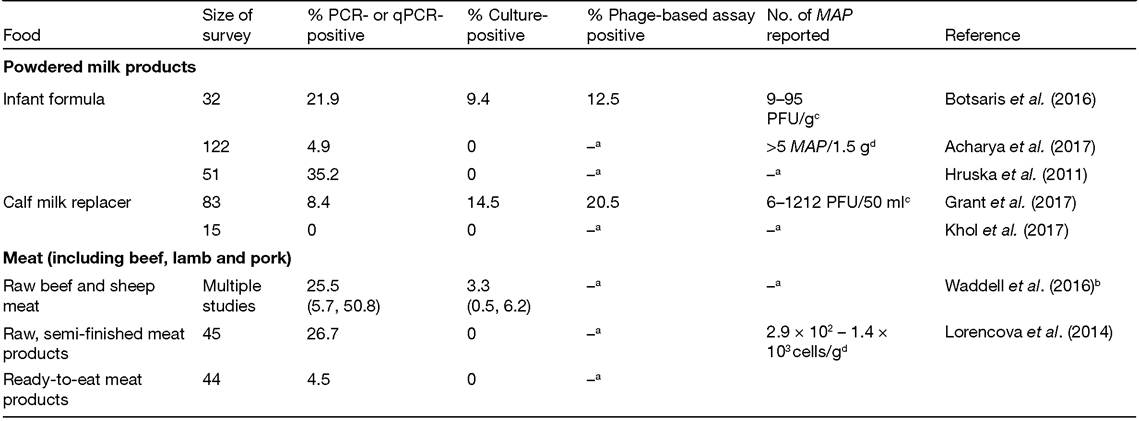
A survey of European Union (EU) infant formula products by Botsaris et al. (2016) reported 9.4% of samples MAP positive by culture and 12.5% by phage-PCR assay, and a survey of US calf milk replacer products by Grant et al. (2017) reported 14.5% of samples MAP positive by culture and 20.5% by PMS-phage assay. Two other surveys of powdered infant formula (Hruska et al., 2011; Acharya et al., 2017) and one of calf milk replacer (Khol et al., 2017) failed to culture viable MAP, although, with the exception of the latter study, there was PCR evidence of the presence of MAP cells in the powdered milk samples tested (Table 2.1).2.2.2 Meat and meat products
MAP contamination of animal carcasses can arise from three sources: (i) faecal contamination on hides being transferred to carcasses during slaughter; (ii) disseminated infection via the bloodstream; and (iii) localized foci of infection, such as lymph nodes, being disrupted, such that the contents are spread to other parts of the carcass or mixed into minced∕ground meat. The risk of MAP contaminating beef is possibly higher when old dairy cows are slaughtered rather than beef cattle, as the latter are generally slaughtered before lesions are evident (Adam and Brulisauer, 2010). However, currently there appear to be no restrictions on paratuberculosis positive cows being sent to slaughter for human consumption, and these animals will pose the greatest risk of MAP transmission via beef, regardless of whether they are from dairy or beef herds. The Waddell et al. (2016) review and meta-analysis considered seven raw beef and sheep studies published up to 2011 and reported a prevalence of 25.5% by PCR methods and 3.3% by culture (Table 2.1). The only MAP and meat survey since that review appears to be Lorencova et al. (2014), who reported the findings of a survey of retail raw, semi-finished and ready-to-eat meat products, comprising minced beef and pork and∕or offal in the Czech Republic. They reported a similar percentage of MAP positivity (26.7%) by qPCR to previous raw meat studies (summarized by Waddell et al., 2016) for raw, semi-finished meat products, but lower qPCR positivity for ready-to-eat meat products (4.5%). Numbers of MAP estimated by qPCR to be present in the retail meat products ranged from 2.9 ? 102 - 1.4 ? 103 cells/g. Viable MAP was not isolated from either type of meat product by culture, but the decontamination method employed (HCl and neutralization with NaOH) may not have been optimal for MAP recovery from meat samples.
Paratuberculosis is also a problem in farmed deer (Power et al., 1993; Fawcett et al., 1995; de Lisle et al., 2003; Kopecna et al., 2008; de Albuquerque et al., 2017) and so there is a theoretical risk of venison being contaminated by MAP in the same way as meat from other infected species. However, no venison surveys have been reported to date. The incidence of MAP in farmed rabbits has been investigated (Arrazuria et al., 2015) because paratuberculosis can also be a problem in rabbits. MAP was not detected in gut- associated lymphoid tissue of 66 rabbits in commercial rabbit farms in Spain by either culture or PCR, but M. avium subsp. avium (10/66, 15.2%) and M. avium subsp. hominissuis (1/66, 1.51%) were, by PCR only.
Generally meat is cooked before consumption and, thus, the effect of cooking on survival of MAP is of interest. A single study, Hammer etal. (2013), has investigated the effect of cooking of hamburger patties made with MAP-spiked beef (at three levels - 102, 104 and 106CFU∕g) for 2-6 min on a hotplate operating at 177°C, with flipping once during cooking. Results in terms of log kill of MAP achieved were ‘highly variable' according to the authors. Cooking to well done, 6 min for 70 g patties achieved a four-log10 reduction, and 5 or 6 min for 50 g patties achieved a more than five-log10 reduction. Internal temperature of the patties was data-logged during cooking, which revealed that not all points in a hamburger achieve the same temperature during cooking. Undercooked, or unevenly cooked, hamburgers may, therefore, represent a food safety risk if high numbers of MAP were present in the raw meat.
2.3
More on the topic Evidence of MAP in Animal- Derived Foods:
- Mycobacterium avium subsp. paratuberculosis in Animal-Derived Foods and the Environment
- Comparative Genotyping of Human and Bovine-Derived MAP Isolates
- NUTRITIONAL PROFILE OF INDIAN FOODS
- Although Nietzsche was referring to slavery in this early essay of his, and not to animal sacrifice, he might well have included sacrifice as a ‘cruel trait', for it involved literally tens of millions of animal victims, many of which did not provide any food.
- Sacrifice: Animals and Other Foods
- The generation of chemical energy by autotrophs, known as primary production, is derived from the uptake of carbon during photosynthesis and chemosynthesis (see Chapter 5).
- Energy that is derived from the consumption of organic compounds produced by other organisms is known as secondary production.
- Comparison of the Virulence and Pathogenicity of MAP Strains
- Genomic Epidemiology of MAP
- MAP Antigens
- Karsten Donat Susanne Eisenberg and Richard WhittingtonThuringian Animal Disease Fund, Animal Health Service, Jena, Germany; 2Justus- Liebig-University, Gieβen, Hesse, Germany; 3Animal Disease Fund of Lower Saxony, Hanover, Germany; 4The University of Sydney, Sydney, Australi
- Pathogenesis and Stages of MAP Infection in Cattle
- Study of MAP Proteins
- Enumeration of MAP
- Cultural Requirements of Different Strains of MAP
- Survival of MAP During Dairy Processing
- Innate Response to MAP Infection
- Genomic Comparison of MAP Strains
- General Principles for the Cultivation of MAP