NPP in aquatic ecosystems is controlled by nutrient availability
The primary producers in lake ecosystems are phytoplankton and rooted macrophytes. NPP in lake ecosystems is often limited by the supply of both phosphorus and nitrogen, as we know not only from the results of experimental manipulations, but also from unintentional “experiments” set in motion by wastewater discharges into lakes (see Figure 22.18).
A common approach to determining the response of NPP in lakes to changes in nutrient supply is to incubate translucent or open-top containers, sometimes referred to as “limnocorrals,” of lake water, amended with one or more nutrients, in the lake (FIGURE 20.14). The NPP response is measured by changes in chlorophyll concentrations or numbers of phytoplankton cells.
FIGURE 20.14 Limnocorrals A researcher snorkels within a contained area (Iimnocorral) in McKinley Lake near Cordova, Alaska, subjected to experimental fertilization to examine the effects of nutrients on NPP. View larger image
One of the most convincing studies of the effect of nutrients on NPP in lakes was a series of whole-lake fertilization experiments by David Schindler (Schindler 1974). The experiments were initiated in 1969 in the Experimental Lakes Area in Ontario, a series of 58 small lakes set aside for experimental manipulations. Concern over declining water quality in the lakes of North America and Europe motivated Schindler and his colleagues to establish several experiments to determine whether inputs of nutrients in wastewater were involved in the dramatic increases in the growth of phytoplankton that had been observed. They added nitrogen, carbon, and phosphorus to all or half of several individual lakes. The results of these experiments provided strong evidence for phosphorus limitation of NPP (FIGURE 20.15). Massive increases in the abundances of cyanobacteria were responsible for the increase in NPP in response to
phosphorus addition.
Evidence for nitrogen limitation of NPP in high-elevation lakes, based on small-scale fertilization experiments and measurements of the ratio of nitrogen to phosphorus in the water, also exists (Elser et al. 2007).
FIGURE 20.15 Response of a Lake to Phosphorus Fertilization Experimental Lake 226 was divided into two sections as part of David Schindler’s experiments on the effects of nutrient availability on NPP. View larger image
NPP in streams and rivers is often low, and the majority of the energy in those ecosystems is derived from terrestrial organic matter (see Concept 21.1). Water movement limits the abundance of phytoplankton, except where the water velocity is relatively low. In Concept 3.2, we introduced the river continuum concept, which describes the increasing importance of in-stream NPP as the river flows downstream. Most of the NPP in streams and rivers comes from photosynthesis by macrophytes and algae attached to the bottom in shallow areas where there is enough light for photosynthesis. Suspended sediment in rivers can limit light penetration; thus, turbidity often controls NPP. Nutrients, particularly nitrogen and phosphorus, can also limit NPP in streams and rivers.
Marine NPP is usually limited by nutrient supply, but the specific limiting nutrients vary among marine ecosystem types. Estuaries, the zones where rivers empty into the ocean (see Concept 3.3), are rich in nutrients relative to other marine ecosystems. Variation in NPP among estuaries is correlated with variation in nitrogen inputs from rivers. Agricultural and industrial activities have increased riverine inputs of nitrogen into estuaries, which have caused periodic “blooms” of algae. These blooms have been implicated in the development of “dead zones”—areas of high fish and zooplankton mortality—in over 400 nearshore ecosystems worldwide.
NPP in the open ocean is derived primarily from phytoplankton, including a group referred to as the picoplankton, consisting of cells smaller than 1 μm.
Picoplankton contribute as much as 50% of the total marine NPP. Smaller contributions come from floating mats of seaweeds such as Sargassum. Near the coast, kelp forests may have leaf area indices and rates of NPP as high as those of tropical forests. “Meadows” of seagrasses such as eelgrass (Zostera spp.) are also important contributors to NPP in shallow nearshore zones.In much of the open ocean, NPP is limited by nitrogen. In the equatorial Pacific Ocean, however, detectable concentrations of nitrogen can be found in the water even when peak NPP occurs, suggesting that some other factor limits NPP. John Martin and colleagues measured the concentrations of nutrients in the open waters of the Pacific and performed bottle incubation experiments with added nutrients. They found that adding iron to the bottles increased NPP (Martin et al. 1994). Based on this evidence that iron limits NPP in some ocean regions,
Martin suggested that windblown dust from Asia, an important source of iron for the open ocean, could play an important role in the global climate system through its influence on marine NPP, and thus on atmospheric CO2 concentrations. During glacial periods, large areas of the continents lacking vegetative cover could have contributed aeolian dust that would have fertilized the ocean. As NPP in marine ecosystems increased, those ecosystems might have taken up more CO2 from the atmosphere, reducing its atmospheric concentration and serving as a positive feedback to cool the climate further. Martin suggested that these findings might be applied to address global warming, saying at the time, “Give me half a tankerload of iron, and I'll give you an Ice Age.” He recommended the use of large-scale experiments to investigate the influence of iron on ocean NPP. Unfortunately, Martin died in 1993, before his ambitious experiments could be carried out.
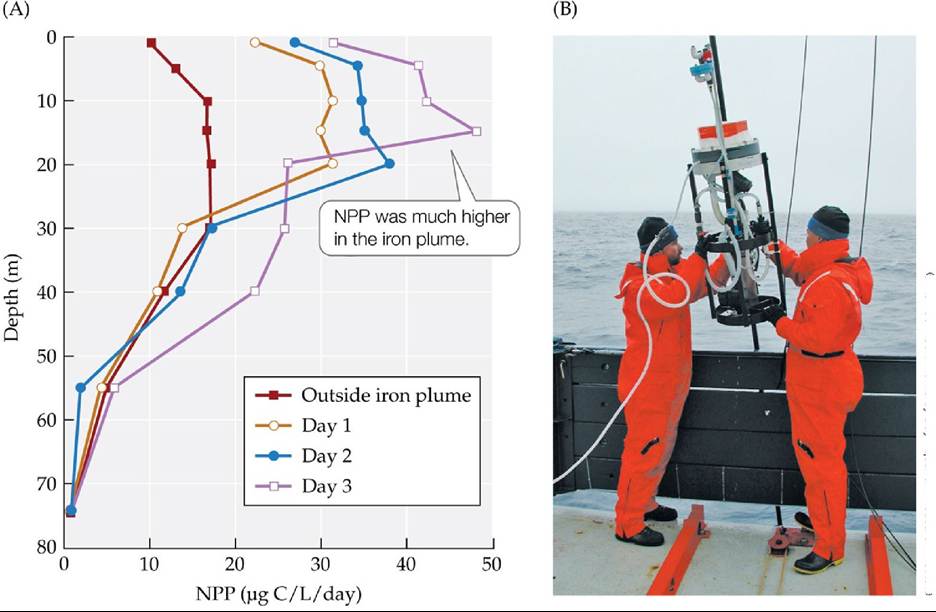
Martin's colleagues subsequently performed the first of several experiments in 1993, adding iron sulfate to surface waters of the equatorial Pacific west of the Galapagos archipelago.
This experiment was alternatively referred to as IronEx I or the “Geritol solution”1 to global climate change. During IronEx I, a 64-km2 area was fertilized with 445 kg of iron, which resulted in a doubling of phytoplankton biomass and a fourfold increase in NPP (FIGURE 20.16). Three other iron fertilization experiments were subsequently performed, one in 1995 (IronEx II), which produced a tenfold increase in phytoplankton biomass; a second in 1999 in the Southern Ocean; and the last in 2002, also in the Southern Ocean. While the iron limitation hypothesis has been strongly supported by these and other experiments, fertilizing large areas of the ocean is unlikely to provide a solution to increasing atmospheric CO2 concentrations and global climate change. Some of the CO2 taken up by phytoplankton is eventually re-emitted to the atmosphere via respiration by zooplankton and bacteria that consume the phytoplankton. In addition, the iron is lost relatively quickly from the surfacephotic zone, sinking to deeper layers where it is unavailable to support phytoplankton photosynthesis and growth. Iron fertilization on a large scale could also have detrimental effects on ocean biodiversity and could create large dead zones similar to those generated by nitrogen inputs into estuaries.
© Ken Johnson/Monterey Bay Aquarium Research Institute
FIGURE 20.16 Effect of Iron Fertilization on Marine NPP IronExlreleasedaplumeof iron into the equatorial Pacific Ocean to study the effects of iron fertilization on NPP. (A) This vertical profile shows primary production at various depths outside and inside the iron plume on three specific days: 1, 2, and 3 days following the release of the iron. (B) Researchers deploy a pump to add iron to the ocean. (A after J. H. Martin et al. 1994. Nature 371: 123-129.) View larger image
The development of remote sensing and eddy covariance techniques has improved our ability to discern global patterns of NPP. We'll examine those patterns in the next section.
1
- Geritol is a dietary supplement once widely believed to help cure “iron-poor, tired blood.”
More on the topic NPP in aquatic ecosystems is controlled by nutrient availability:
- NPP in terrestrial ecosystems is controlled by climate
- Nutrients in Aquatic Ecosystems
- CONCEPT 22.4 Freshwater and marine nutrient cycles occur in a moving medium and are linked to terrestrial ecosystems.
- NPP changes during ecosystem development
- Terrestrial and oceanic NPP are nearly equal
- NPP can be estimated by a number of methods
- Differences among biomes in NPP reflect climate and biotic variation
- MORBILLIVIRUS INFECTIONS IN AQUATIC MAMMALS
- Seasonal changes in aquatic environments are associated with changes in water temperature and density
- Decomposition is a key nutrient recycling process
- Organisms have specific nutrient requirements
- INFLUENZA IN AQUATIC MAMMALS
- Which biomes and marine biological zones have the highest NPP and, as a consequence, the greatest effect on atmospheric CO2 dynamics?
- MYCOPLASMA INFECTIONS OF AQUATIC MAMMALS
- Herpesvirus infections in AQUATIC MAMMALS