Organisms influence their temperature by modifying energy balance
On a hot day, jumping into a swimming pool and then sitting in the shade in a light breeze brings relief from the oppressive heat. Elephants follow a similar routine, wading into ponds and using their trunks to spray water onto their backs.
This kind of behavior facilitates heat loss in several ways. First, the contact of warm skin with cool water causes heat energy to be lost from the body through the process of conduction: the direct transfer of energy from warmer, more rapidly moving molecules to cooler, more slowly moving molecules. Also, when cool water and air move across the surface of a warmer body, heat energy is carried away via convection. In addition, the change in the state of water from liquid to vapor as it evaporates on the skin's surface absorbs body heat (latent heat transfer). Finally, moving into the shade lowers the amount of energy you receive from solar radiation.The balance between energy input and energy output determines whether the temperature of any object will increase or decrease. Archaea, bacteria, fungi, protists, and algae cannot avoid changes in their temperature when the environmental temperature changes. They must tolerate variations in temperature through biochemical modifications. For example, when temperatures exceed their range of tolerance, microorganisms often survive as dormant spores. Plants and animals can influence their body temperature, and therefore their physiological processes, by adjusting their exchange of energy with the environment. Both plants and animals are often able to avoid stressful internal temperatures through behavioral and morphological modifications of energy balance. Let's examine some examples.
Modification of Energy Balance by Plants
Among plants, temperature stress is experienced mainly in terrestrial environments. Marine and aquatic plants usually experience temperatures within the range that is conducive to their physiological functioning, although those in nearshore habitats can experience potentially lethal temperatures.
The factors involved in the energy balance of terrestrial plants are shown in FIGURE 4.8. Energy inputs that warm the plant include sunlight and infrared radiation from surrounding objects. If the ground or air is warmer than the plant, energy inputs also include conduction and convection. Losses of energy from the plant include the emission of infrared radiation to the surrounding environment and, if the ground or air is cooler than the plant, conduction and convection. Heat loss also occurs through transpiration (evaporation of water from inside the plant) andsurface evaporation, collectively referred to as evapotranspiration.
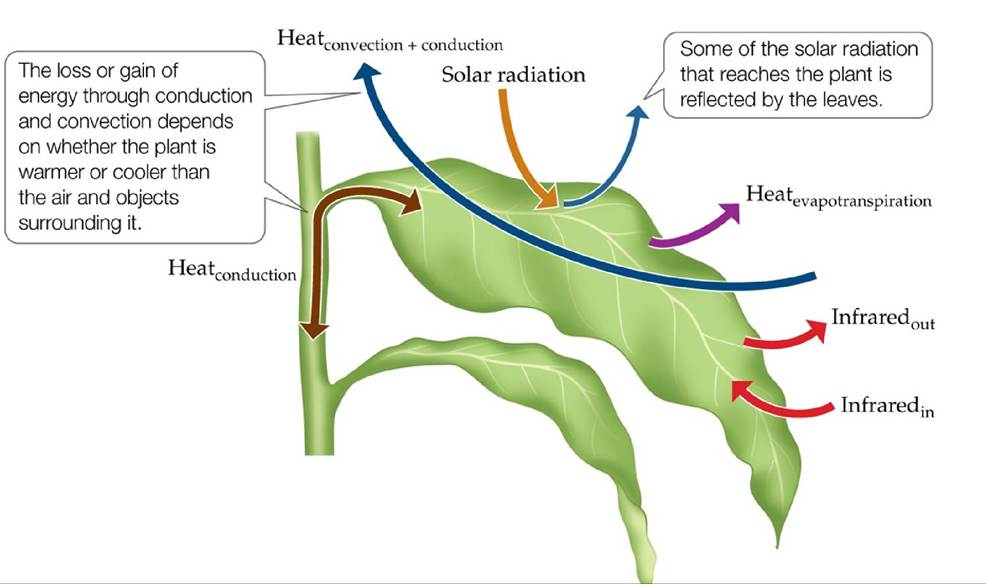
FIGURE 4.8 EnergyExchangeinTerrestrialPlants Thetemperatureofaplantis determined by the balance between inputs of energy from and outputs of energy to the environment. (After P. S. Nobel. 1983. Biophysical Plant Physiology and Ecology. W. H. Freeman: New York.) View larger image
We can put these inputs and outputs together to determine whether the temperature of the plant is changing:

(4.1)
where ΔHplant is the heat energy change of the plant (the Greek delta usually signifies “change in”), SR is solar radiation, IRin is the input of infrared radiation, IRout is the output of infrared radiation, Hconv is convective heat transfer, Hcond is conductive heat transfer, and Het is heat transfer through evapotranspiration. A negligible loss of energy occurs as the plant uses solar radiation for photosynthesis. If the plant is warmer than the surrounding air, Hconv and Hcond are negative. If the sum of the energy inputs exceeds the sum of the outputs, ΔHplant is positive, and the plant's temperature is increasing. Conversely, if more heat is being lost than gained, ΔHplant is negative, and the plant's temperature is decreasing.
Plants can modify their energy balance to control their temperature by adjusting these energy inputs and outputs. Leaves are most often associated with these adjustments because they are the primary photosynthetic organs of the plant and typically are the most temperature-sensitive tissue. The most important and common adjustments include changes in the rate of transpirational water loss. In addition, changes in leaf surface reflective properties (color) or in leaf orientation toward the sun can alter the amount of solar radiation absorbed by the plant. Finally, changes in convective heat transfer can be accomplished by changing surface roughness.
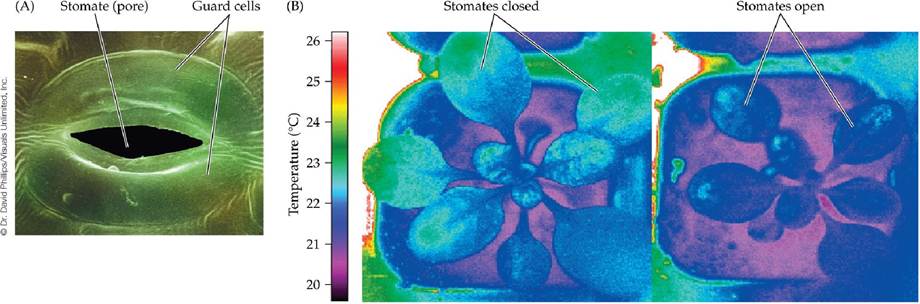
Transpiration is an important evaporative cooling mechanism for leaves. As we saw in Chapter 2, its effectiveness is especially evident in the canopies of tropical forests, which are subjected to warm air temperatures and high levels of solar radiation. Without transpirational cooling, the leaves of tropical canopy plants could reach temperatures over 45°C (>113°F), which would be lethal. The rate of transpiration is controlled by specialized guard cells surrounding pores, called stomates, leading to the interior of the leaf. Stomates are the gateway for both transpirational water loss and the uptake of carbon dioxide for photosynthesis; we will return to the latter function in Concept 5.2. Variation in the degree of stomatal opening, as well as in the number of stomates, controls the rate of transpiration and therefore exerts an important control on leaf temperature (FIGURE 4.9).
Courtesy of G. H. Holroyd and A. t√1. Heatherington
FIGURE 4.9 Stomates Control Leaf Temperature by Controlling Transpiration (A) Specialized guard cells control a stomate's degree of opening. Open stomates allow CO2 to diffuse in for photosynthesis, and they allow water to transpire out, cooling the leaves. (B) Leaf temperatures vary with the degree of stomatal opening.
The plant on the right has open stomates and is transpiring freely, while the plant on the left, kept under identical conditions, has closed stomates, a lower transpiration rate, and a temperature 1°C-2°C (2°F-4°F) higher, as indicated by thermal infrared imaging.Cooling of leaves using transpiration may be particularly important in what biomes?
View larger image
Transpiration requires a steady supply of water. Where the amount of water in the soil is limited—as it is over a substantial part of Earth's land surface— transpiration is not a reliable cooling mechanism. As we saw in Concept 3.1, some plants shed their leaves during dry seasons, thereby avoiding both temperature and water stress. However, the high demand for the resources (e.g., soil nutrients) needed to replace fallen leaves may favor protecting existing leaves rather than shedding them. Plants that maintain their leaves during long dry periods require mechanisms other than transpiration to dissipate heat energy. One option is to alter the reflective properties of leaves via pubescence, the presence of light-colored or white hairs on the leaf surface, which lowers the amount of solar radiation absorbed by the leaf surface. Pubescence can also lower the effectiveness of convective heat loss, however, and thus represents a trade-off between two opposing heat exchange mechanisms.
One of the best studies addressing the adaptive significance of leaf pubescence for temperature regulation has focused on shrubs of the genus Encelia (members of the daisy family, common name brittlebush). Jim Ehleringer and his colleagues described the role of pubescence in leaf temperature regulation among species of Encelia that occupy different geographic ranges. Encelia farinosa, a native of the Sonoran and Mojave Deserts, maintains a high amount of leaf pubescence relative to Encelia shrubs from moister, cooler environments. Ehleringer and his colleague Craig Cook (1990) evaluated the relative roles of leaf pubescence and transpiration in the cooling of leaves of E.
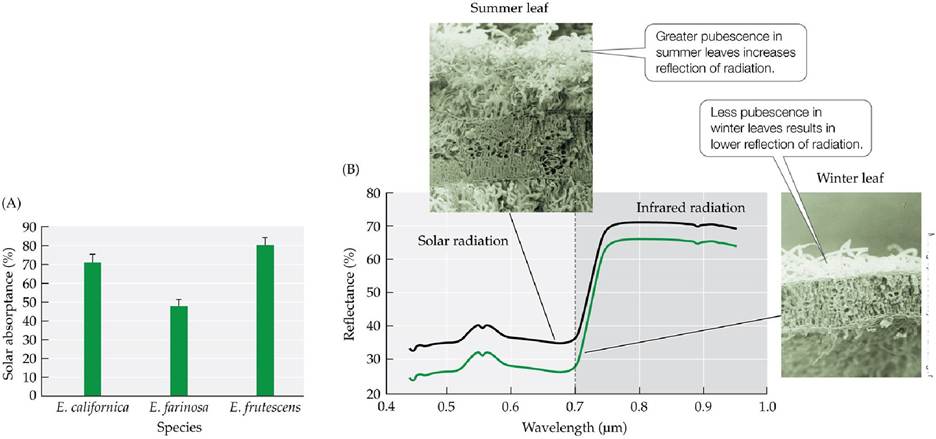
farinosa and two other species whose leaves lack pubescence: E. frutescens, which occurs in desert washes (which have more moisture than the rest of the desert), and E. californica, native to the cooler, moister coastal sage community of California and Baja California. To control for environmental variation that could influence the morphology and physiology of the plants, they grew plants of each species from seed together in experimental plots in the Sonoran Desert and on the California coast. Half of their experimental plants were watered while the other half were left under natural conditions. They measured the leaf temperatures, the degree of stomatal opening, and the amount of sunlight absorbed.The three Encelia species showed few differences in leaf temperature and stomatal opening when grown in the cooler, moister California coastal garden. In the desert garden, however, E. californica and E. frutescens shed their leaves during the hot summer months under natural conditions, but E. farinosa did not. Encelia frutescens did not shed its leaves when the shrubs were watered, and its leaves maintained sublethal temperatures using transpirational cooling. Encelia farinosa leaves reflected about twice as much solar radiation as those of the other two species (FIGURE 4.10A), which facilitated the shrub's ability to maintain leaf temperatures lower than the air temperature.
Micrographs courtesy of J. Ehleringer
FIGURE 4.10 Sunlight, Seasonal Changes, and Leaf Pubescence (A)Solarheatingof leaves varies according to the amount of pubescence on those leaves. The pubescent leaves of the desert shrub Encelia farinosa absorb a lower percentage of the incoming solar radiation than the leaves of two nonpubescent species: E. californica, native to the coastal sage community of California, and E. frutescens, an inhabitant of moister desert wash communities. Encelia farinosa is therefore less dependent on transpiration for leaf cooling than the other two species.
Error bars show 1 standard error of the mean. (B) Encelia farinosa produces greater amounts of pubescence on its leaves in summer than in winter, representing acclimatization to hot summer temperatures. The photos are scanning electron micrographs of leaf cross sections.Why might temperature regulation associated with greater reflection of solar radiation via pubescence be more important in deserts than in a warm, moist biome such as the tropical rainforest?
(A after J. R. Ehleringer and C. S. Cook. 1990. Oecologia 82: 484-489.) View larger image
Ehleringer and Cook's field experiment provides correlative evidence of the adaptive value of leaf pubescence to E. farinosa under hot desert conditions. Additional work by Darren Sandquist and Ehleringer has supported its adaptive value, indicating that natural selection has acted on variation in pubescence among ecotypes of E. farinosa. Populations in drier environments have more leaf pubescence, and reflect more solar radiation, than populations from moister environments (Sandquist and Ehleringer 2003).
In addition to varying among species and populations, leaf pubescence can also vary seasonally, exemplifying acclimatization to environmental conditions. Encelia farinosa shrubs produce smaller, more pubescent leaves in summer and larger, less pubescent leaves in winter (FIGURE 4.10B). There are costs to being pubescent, associated with the construction of the hairs and the loss of solar radiation that could be used for photosynthesis. Thus, when temperatures are cooler or when reliable soil water is present, E. farinosa plants construct leaves with fewer hairs.
Heat can be lost from a leaf by convection when the air temperature is lower than the temperature of the leaf. The effectiveness of convective heat loss is related to the speed of the air moving across a surface. As the moving air experiences more friction closer to the surface of an object, the flow becomes more turbulent, forming eddies (FIGURE 4.11). This zone of turbulent flow, called the boundary layer, lowers convective heat loss. The thickness of the boundary layer around a leaf is related to its size and its surface roughness. Small, smooth leaves have thin boundary layers and lose heat more effectively than large or rough leaves. This relationship between the boundary layer and convective heat loss is one reason for the rarity of large leaves in desert ecosystems.
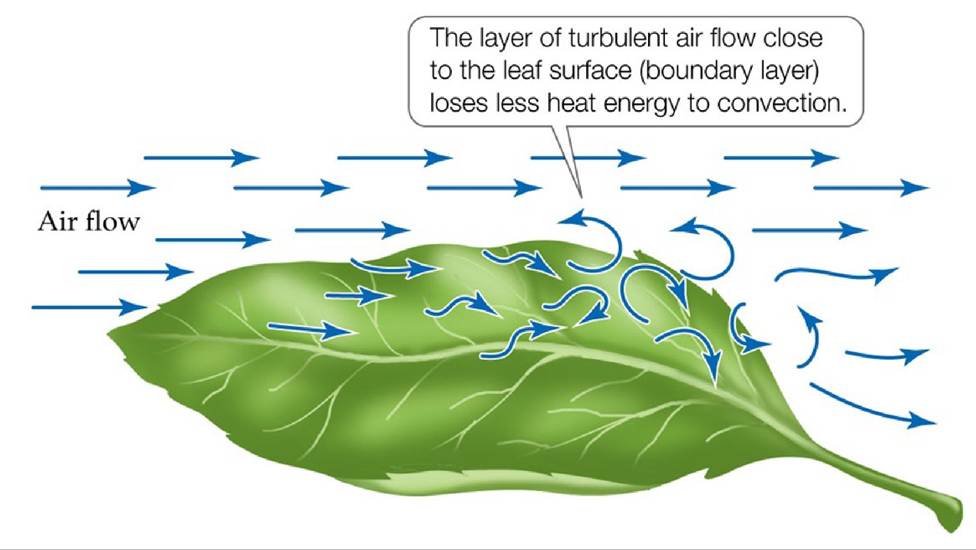
FIGURE 4.11 ALeafBoundaryLayer Air flowing close to the surface of a leaf is subject to friction, which causes the flow to become turbulent and lowers convective heat loss from the leaf to the surrounding air. View larger image
Excessive heat loss by convection can be a problem for plants (and animals) in cold, windy environments such as the alpine zone in mountains. Convection is the largest source of heat loss from the land surface in temperate alpine environments, and high winds can shred leaves in exposed sites. Most alpine plants grow close to the ground surface to avoid the high wind velocities. Some alpine plants produce a layer of insulating hair on their surface to lower convective heat loss. The snow lotus of the Himalayas (Saussurea medusa) produces a series of very densely pubescent leaves that surround the flowers of the plant (FIGURE 4.12). Although they project above the ground surface and are exposed to more wind than ground-hugging plants, the flowers of S. medusa remain as much as 20°C (36°F) warmer than the air by absorbing and retaining solar radiation (Tsukaya et al. 2002). The plant not only keeps its photosynthetic tissues warm, but also provides a warm environment for potential pollinators,
which are in short supply in cold, windy alpine environments.

FIGURE 4.12 A Woolly Plant of the Himalayas The snow lotus (Saussurea medusa) has dense pubescence surrounding its emergent flowering stems, which provides them with thermal insulation. View larger image
Modification of Energy Balance by Animals
Animals are subject to the same energy inputs and outputs described for plants in Equation 4.1, with one key difference: some animals—in particular, birds and mammals—have the ability to generate heat internally. As a result, another term is needed in the energy balance equation to represent this metabolic heat generation:
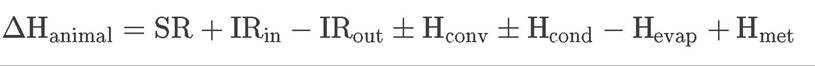
(4.2)
where ΔHanimal is the heat energy change of the animal, SR is solar radiation, IRin is the input of infrared radiation, IRout is the output of infrared radiation, Hconv is convective heat transfer, Hcond is conductive heat transfer, Hevap is heat transfer through evaporation, and Hmet is metabolic heat generation. In contrast to plants, evaporative heat loss is not widespread among animals. Notable examples of evaporative cooling in animals include sweating in humans, panting by dogs and other animals, and licking of the body by some marsupials under conditions of extreme heat.
The internal generation of heat by some animals represents a major ecological advance. Animals capable of metabolic heat generation can maintain relatively constant internal temperatures near the optimum for physiological functioning under a wide range of external temperatures, and as a result, they can expand their geographic ranges. Varying degrees of reliance on internal heat generation exist throughout the animal kingdom. Animals that regulate their body temperature primarily through energy exchange with the external environment, which includes the majority of animal species, are called ectotherms. Animals that rely primarily on internal heat generation, which are called endotherms, include, but are not limited to, birds and mammals. Internal heat generation is also found in some fishes (e.g., tuna), insects (e.g., bees, which generate heat for metabolic function as well as for defense; FIGURE 4.13), and even a few plant species (e.g., skunk cabbage, Symplocarpus foetidus, which warms its flowers using metabolically generated heat during the spring).

FIGURE 4.13 Internal Heat Generation as a Defense Beescangenerateheatby contracting their flight muscles. Japanese honeybees (Apis cerana) use internal heat generation as a defense against Asian giant hornets (Vespa mandarinia) that attack bee colonies. (A) When a hornet enters a nest, the honeybees swarm the larger invader. (B) The defensive ball of bees surrounding an invading hornet generates enough heat that temperatures in the center exceed the upper lethal temperature for the hornet (about 47°C, 117°F), thus killing the invader. View larger image
Temperature Regulation and Tolerance in Ectotherms
Generally, ectotherms have a greater tolerance for variation in their body temperature than endotherms (see Figure 4.7), possibly because they are less able to adjust their body temperature than endotherms. The exchange of heat between an animal and the environment, whether for cooling or heating, depends on the amount of surface area relative to the volume of the animal. A larger surface area relative to volume allows greater heat exchange but makes it harder to maintain a constant internal temperature in the face of variable external temperatures. A smaller surface area relative to volume decreases the animal's ability to gain or lose heat. This relationship between surface area and volume imposes a constraint on the body size and shape of ectothermic animals. Generally speaking, the surface-area-to-volume ratio decreases as body size increases, and the animal's ability to exchange heat with the environment decreases as well. As a result, large ectothermic animals are considered improbable.
Small aquatic ectotherms (e.g., most invertebrates and fishes) generally remain at the same temperature as the surrounding water. Some larger aquatic animals, however, can maintain a body temperature warmer than that of the surrounding water (FIGURE 4.14). For example, skipjack tuna (Katsuwonus pelamis) use muscle activity, in conjunction with heat exchange between blood vessels, to maintain a body temperature as much as 14°C (25°F) warmer than the surrounding seawater. Other large oceanic fishes use similar circulatory heat exchange mechanisms to keep their muscles warm. Such mechanisms are particularly important for predatory species that depend on rapid acceleration to capture prey, which is aided by having warmer muscles.
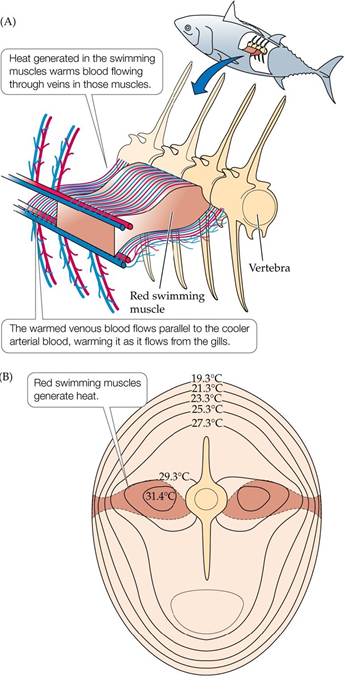
FIGURE 4.14 Internal Heat Generation by Tuna (A) Heat generated in the red swimming muscles of the skipjack tuna, used for cruising through the water, warms blood flowing through them, which is carried toward the body surface in veins. Those veins run parallel to arteries carrying cool oxygenated blood from the gills, warming that blood before it reaches the swimming muscles. (B) A cross section of the tuna shows that its core remains warmer than the surrounding water. View larger image
The mobility of many terrestrial ectotherms allows them to adjust their body temperature by moving to places that are warmer or cooler than they are. Basking in the sun or moving into the shade allows these animals to adjust their energy gains and losses via solar radiation, conduction, and infrared radiation. For example, reptiles and insects emerging from hiding places after a cool night will bask in the sun to warm their bodies prior to initiating their daily activities (FIGURE 4.15). This basking behavior, however, increases their risk of being found by predators. Many of these animals rely on camouflage (also called
crypsis) to escape detection while basking. In addition to moving between locations with different temperatures, reptiles may regulate their body temperatures by altering their coloration and changing their orientation to the sun.
111" class="lazyload" data-src="/files/uch_group80/uch_pgroup312/uch_uch7303/image/image110.jpg">
FIGURE 4.15 Mobile Animals Can Use Behavior to Adjust Body Temperature Anadult female saltwater crocodile (Crocodylus porosus) sunning itself on the riverbank, Daintree River, Daintree National Park, Far North Queensland.
What components of energy exchange are affected by the behavior of this crocodile?
View larger image
Because they rely on the external environment for temperature regulation, the activities of ectothermic animals are limited to certain temperature ranges. When temperatures are warm, ectotherms in sunny environments (e.g., deserts) may gain enough energy from the environment to push their body temperatures to lethal levels. Increases in temperature associated with climate change over the past two decades appear to have limited the daily foraging periods of several species of Mexican lizards, whose abundances have decreased significantly
during this period (also see Concept 25.2).
In temperate and polar regions, temperatures drop below freezing for extended periods. Ectotherms inhabiting these regions must either avoid or tolerate exposure to subfreezing temperatures. Avoidance may take the form of seasonal migration (e.g., moving to a lower latitude) or movement to local microhabitats where temperatures stay at or above freezing (e.g., burrowing into the soil). Tolerance of subfreezing temperatures involves minimizing the damage associated with ice formation in cells and tissues. If ice forms as crystals, it will puncture cell membranes and disrupt metabolic functioning. Some insects inhabiting cold climates contain high concentrations of glycerol, a chemical compound that minimizes the formation of ice crystals and lowers the freezing point of body fluids. These insects spend winter in a semifrozen state, emerging in spring when temperatures are more conducive to physiological activity. Vertebrate ectotherms generally do not tolerate freezing to the degree that invertebrate ectotherms do, because of their larger size and greater physiological complexity. A very few amphibians, however, can survive being partially frozen, as described in the Case Study at the opening of this chapter.
Temperature Regulation and Tolerance in Endotherms
Endotherms tolerate a narrower range of body temperatures (30oC-45oC, 86°F- 113°F) than ectotherms. However, the ability of endotherms to generate heat internally has allowed them to greatly expand their geographic ranges and the times of year they can be active. Endotherms can remain active at subfreezing environmental temperatures, something that most ectotherms cannot do. The cost of being endothermic is a high demand for food to supply energy to support metabolic heat production. The rate of metabolic activity in endotherms is associated with the external temperature and the rate of heat loss. The rate of heat loss, in turn, is related to body size because of its influence on surface-area- to-volume ratio. Small endotherms have higher metabolic rates, require more energy, and have higher feeding rates than large endotherms.
Endothermic animals maintain a constant basal (resting) metabolic rate over a range of environmental temperatures known as the thermoneutral zone. Within the thermoneutral zone, minor behavioral or morphological adjustments are sufficient for maintaining an optimal body temperature. When the environmental temperature drops to a point at which heat loss is greater than metabolic heat production, the body temperature begins to drop, triggering an increase in metabolic heat generation. This point is called the lower critical temperature (FIGURE 4.16A). The thermoneutral zone and the lower critical temperature differ among mammal species (FIGURE 4.16B). As one would expect, mammals from the Arctic have lower critical temperatures below those of animals from tropical regions. Note also that the rate of metabolic activity (slope of the line) increases more rapidly below the lower critical temperature in tropical than in Arctic mammals.
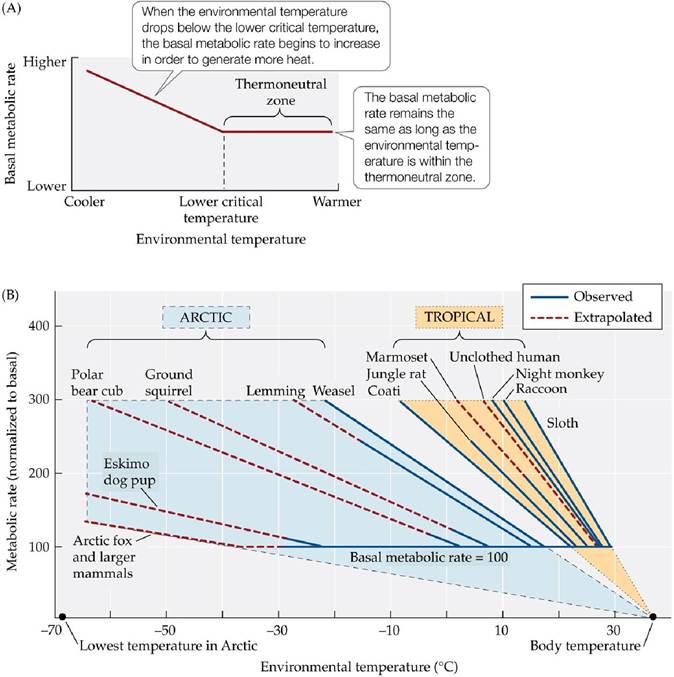
FIGURE 4.16 Metabolic Rates in Endotherms Vary with Environmental Temperatures
(A) An endotherm’s resting, or basal, metabolic rate stays constant throughout a range of environmental temperatures known as the thermoneutral zone. When environmental temperatures reach a lower limit, known as the lower critical temperature, the endotherm’s metabolic rate increases to generate additional heat. (B) The thermoneutral zones and lower critical temperatures of endotherms vary with their habitats. The lower critical temperatures of Arctic endotherms are lower than those of tropical endotherms, and their metabolic rates increase more slowly below those lower critical temperatures, as shown by the shallower slopes of the curves. (B after P. F. Scholander et al. 1950. Biol Bull 99: 237-258.) View larger image
What causes these differences in metabolic adjustments between endotherms of different biomes? For endothermy to work efficiently, animals must be able to retain their metabolically generated heat. Thus, the evolution of endothermy in birds and mammals required insulation: feathers, fur, or fat. These insulating layers provide a barrier limiting conductive (and, in some cases, convective) heat loss. Fur and feathers insulate primarily by providing a layer of still air, similar to a plant's boundary layer, adjacent to the skin. Differences in insulation help explain the differences among the endotherms in Figure 4.16B. Arctic mammals generally maintain thick fur. In warmer climates, however, the ability to cool off through conduction and convection is inhibited by insulation, and thick fur can be an impediment to maintaining an optimal body temperature. Some endotherms acclimatize to seasonal temperature changes by growing thicker fur in winter and shedding fur when temperatures get warmer (a fact that most pet owners know well) (ANALYZING DATA 4.1). Our human ancestors evolved in the hot tropical regions of Africa and lost much of their hairy insulating layer about 2 million years ago (Jablonski 2006).
Z ∖
ANALYZING DATA 4.1
How Does Fur Thickness Influence Metabolic Activity in Endotherms?
Some endotherms exhibit seasonal changes in fur thickness, which helps to enhance heat loss during summer and retain heat generated by the body during winter. This seasonal change in fur thickness in individual animals is an example of acclimatization to changes in temperature.
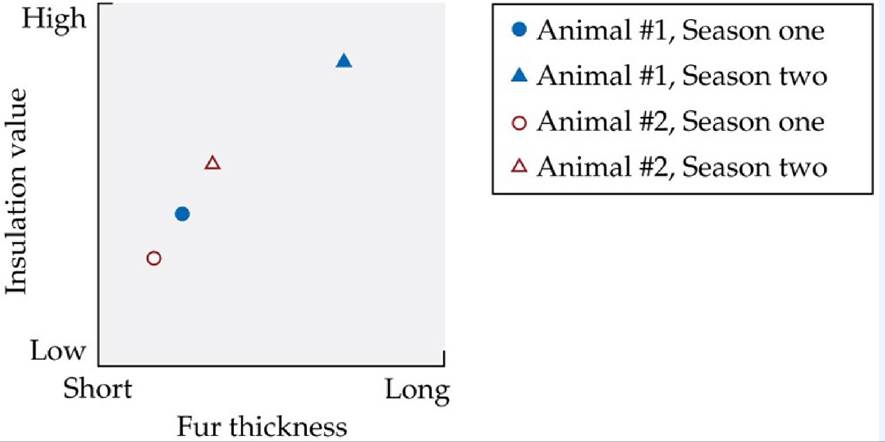
The graph* shows the insulation value (how well heat is retained) versus fur thickness for two animals of the boreal forest biome, a red squirrel (Tamiasciurus hudsonicus) and a wolf (Canis lupus). Both animals are endotherms that exhibit acclimatization to seasonal temperature changes by changes in fur thickness.
1. Each animal is represented by fill or no-fill. Which do you think belongs to which animal (circles and triangles), and why?
2. Which season (summer or winter) is represented by the circles, and which by the triangles? Which animal experiences greater seasonal acclimatory changes in fur length? In what additional way might the animal with the smaller change in fur length cope with extreme winter cold?
*After P. Willmer et al. 2005. Environmental Physiology of Animals, 2nd ed. Blackwell Publishing: Malden, MA.
Cold climates are tough on small endotherms. Small mammals, by necessity, have thin fur, since thick fur would inhibit their mobility. The high demand for metabolic energy below the lower critical temperature, the low insulation values of their fur, and their low capacity to store energy make small mammals improbable residents of polar, alpine, and temperate habitats. However, the faunas of many of these cold climates contain many species of small endotherms, sometimes in high abundances. What explains this apparent discrepancy? Small endotherms, such as rodents and hummingbirds, are able to alter their lower critical temperature during cold periods by entering a state of dormancy known as torpor. The body temperatures of animals in torpor may drop as much as 20°C (36°F) below their normal temperatures. The metabolic rate of an animal in torpor is 50%-90% lower than its basal metabolic rate, providing substantial energy savings (Schmidt-Nielsen 1997). However, energy is still needed to arouse the animal from torpor and bring its body temperature back up to its usual set point. Thus, the length of time an animal can remain in torpor is limited by its reserves of energy. Small endotherms may undergo daily torpor to minimize the energy needed during cold nights. Torpor lasting several weeks during the winter, sometimes referred to as hibernation, is possible only for animals that have access to enough food and can store enough energy reserves, such as marmots (FIGURE 4.17). Hibernation is somewhat rare in polar climates because few animals have access to enough food to provide enough stored energy (in the form of fat) to get through winter without eating. Some large animals, such as bears, enter a long-term winter sleep (sometimes called denning) during which the body temperature decreases only slightly, rather than going into torpor.
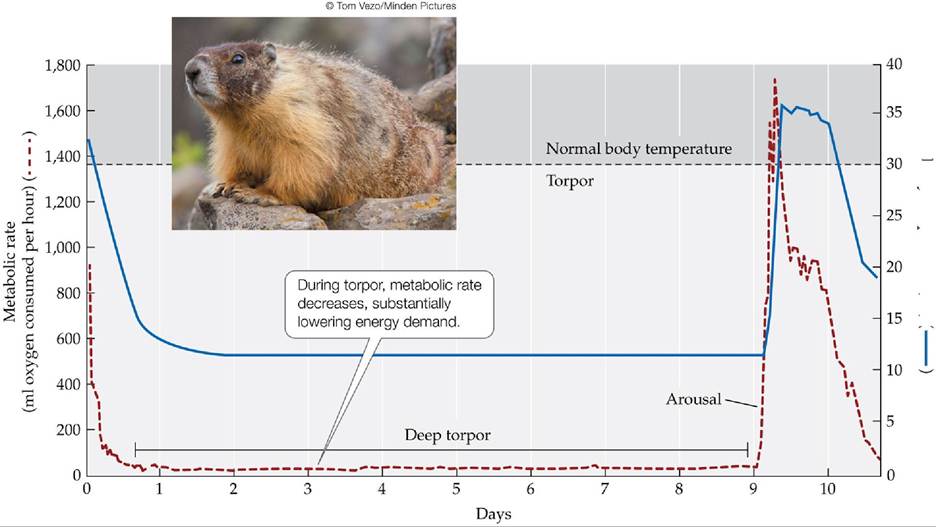
Body temperature (0C) (■
FIGURE 4.17 Long-TermTorporinMarmots Torpor allows yellow-bellied marmots
(Marmota flaviventris) to conserve energy during winter, when food is scarce and the demand for metabolic energy to keep warm is high. Regular cycles of arousal and return to torpor occur for unknown reasons. (After K. Armitage et al. 2003. CompBiochemPhys 134A: 101-114.) View larger image
Just as organisms must balance energy input and output to maintain an optimal temperature, they must balance the movement of water into and out of their bodies to maintain optimal conditions for physiological functioning.
More on the topic Organisms influence their temperature by modifying energy balance:
- CONCEPT 21.3 Changes in the abundances of organisms at one trophic level can influence energy flow at multiple trophic levels.
- Global energy balance drives the climate system
- Mountains influence wind patterns and gradients in temperature and precipitation
- CONCEPT 4.2 The temperature of an organism is determined by exchanges of energy with the external environment.
- Energy that is derived from the consumption of organic compounds produced by other organisms is known as secondary production.
- The second law of thermodynamics states that during any transfer of energy, some energy is dispersed as unusable energy because of the tendency toward an increase in disorder (entropy).
- CONCEPT 5.1 Organisms obtain energy from sunlight, from inorganic chemical compounds, or through the consumption of organic compounds.
- Explain how interactions between organisms and their environment can affect other organisms and potentially lead to unexpected consequences.
- Energy flow between trophic levels can be depicted using energy or biomass pyramids
- The vast majority of the autotrophic production of chemical energy on Earth occurs through photosynthesis, a process that uses sunlight to provide the energy needed to take up carbon dioxide and synthesize organic compounds, principally carbohydrates.
- Disease-Modifying Antirheumatic Drugs
- CONCEPT 5.2 Radiant and chemical energy captured by autotrophs is converted into stored energy in carbon-carbon bonds.
- Temperature
- Temperature controls physiological activity
- Primary abiotic factors and adaptations of organisms
- Working independently of each other, A. J. Lotka (1932) and Vito Volterra (1926) both modeled competition by modifying the logistic equation.
- Climate controls where and how organisms live
- Seasonal changes in aquatic environments are associated with changes in water temperature and density
- High Environmental Temperature
- Radiation Balance of Earth