Water losses and solute gains and losses must be compensated
Terrestrial plants and soil microorganisms rely on water uptake from soils to replace the water they lose to the atmosphere. Soils are important reservoirs of water that support a multitude of ecological functions.
The amount of water that soils can store is related to the balance between water inputs and outputs, soil texture, and topography (FIGURE 4.18). Water inputs include precipitation that infiltrates into the soil and overland flow of water. Water losses include percolation to deeper layers below the plant rooting zone and evapotranspiration.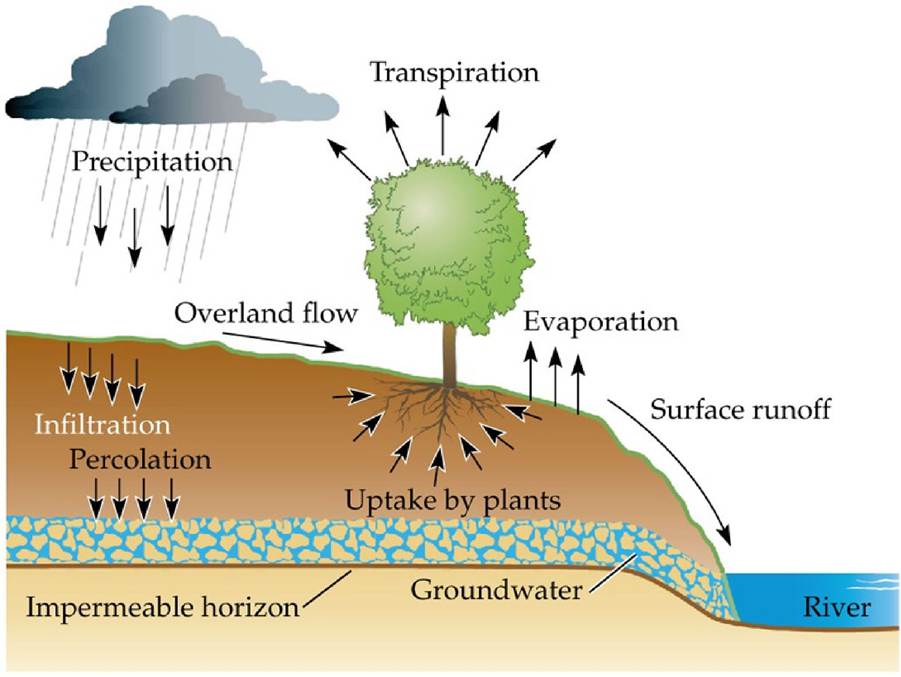
FIGURE 4.18 What Determines the Water Content of Soil? Thewatercontentofsoilis determined by the balance between water inputs (infiltration of precipitation and overland flow of water) and outputs (percolation to deeper layers, evapotranspiration) and by the capacity of the soil to hold water. Soil water storage capacity and the rate of percolation are dependent on soil texture. (After P. J. Kramer. 1983. Water Relations of Plants. Academic Press: Cambridge, MA.) View larger image
The water storage capacity of most soils is dominated by their pore space and matric potential, which is related to the attractive forces on the surfaces of the soil particles. Sandy soils store less water than fine-textured soils, but fine soil particles also have a higher matric potential and thus hold on to water more tightly. Soils with mixed coarse and fine particles are generally most effective in storing water and supplying it to plants and soil organisms. When the volume of water in the soil drops below a certain point (25% of total soil mass in fine- textured soils, 5% in sandy soils), the matric forces are strong enough that most of the remaining water is unavailable to organisms. The osmotic potential of some soils also can be important, particularly where dissolved salts are found, as in soils near marine environments or where salinization (see Figure 2.25) has occurred.
Water Balance in Microorganisms
Single-celled microorganisms, which include archaea, bacteria, algae, and protists, are active primarily in aqueous environments. Their water balance is dependent on the water potential of the surrounding environment, which is determined mainly by its osmotic potential. In most marine and freshwater ecosystems, the osmotic potential of the environment changes little over time. Some environments, however, such as estuaries, tide pools, saline lakes, and soils, experience frequent changes in osmotic potential due to evaporation or variable influxes of fresh and salt water. Microorganisms in these environments must respond to these changes by altering their cellular osmotic potential if they are to maintain a water balance suitable for physiological functioning. They accomplish this through osmotic adjustment, an acclimatization response that involves changing their solute concentration, and thus their osmotic potential. Some microorganisms synthesize organic solutes to adjust their osmotic potential, which also helps to stabilize enzymes. Others use inorganic salts from the surrounding medium for osmotic adjustment. The ability to adjust osmotic potential in response to changes in external water potential varies substantially among microorganisms: some completely lack this ability, while others (such as Halobacterium spp.) can adjust to even the extremely saline conditions in landlocked saline lakes.
As noted above, terrestrial environments are too dry for any organism that is unable to restrict cellular water loss to the atmosphere. Many microorganisms avoid exposure to dry conditions by forming dormant resistant spores, encasing themselves in a protective coating that prevents water loss to the environment. Some microorganisms with filamentous forms, such as fungi and yeasts, are very tolerant of low water potentials and can grow in dry environments. Most terrestrial microorganisms, however, are found in soils, which have a higher water content and humidity than the air above them.
Water Balance in Plants
One of the distinguishing characteristics of plants is a rigid cell wall composed of cellulose. Bacteria and fungi also have cell walls, composed of materials such as chitin (in fungi) or peptidoglycans and lipopolysaccharides (in bacteria). Cell walls are important to water balance because they facilitate the development of positive turgor pressure. When water follows a gradient of water potential into a plant cell, it causes the cell to expand and press against the cell wall, which resists the pressure because of its rigidity (FIGURE 4.19). Turgor pressure is an important structural component of plants, and it is also an important force for growth, promoting cell division. When nonwoody plants lose turgor pressure due to dehydration, they wilt. Wilting is generally a sign that a plant is experiencing water stress.
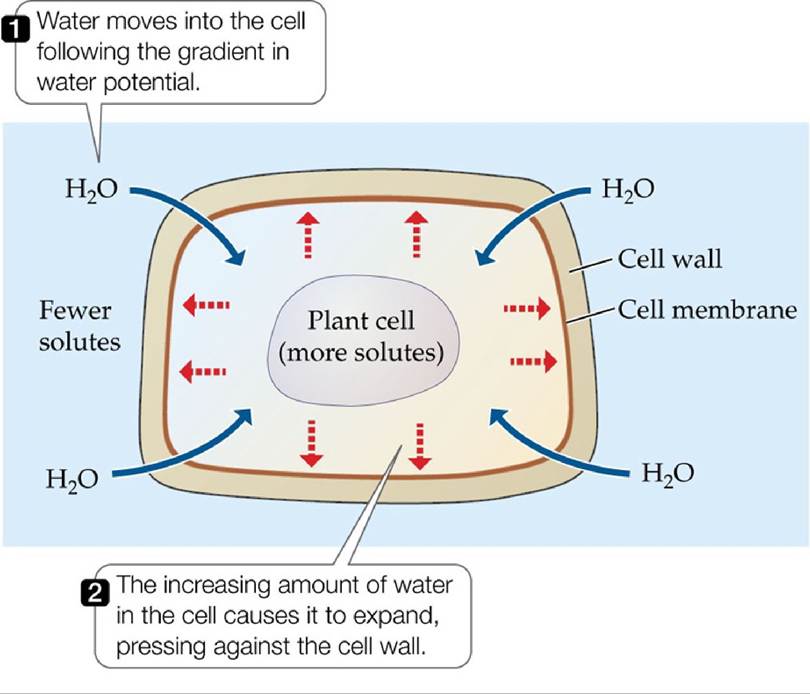
FIGURE 4.19 Turgor Pressure in Plant Cells When a plant cell is surrounded by water with a solute concentration lower than its own, water moves into the cell, while solutes in the cell are prevented from moving out by the cell membrane. The increasing amount of water in the cell causes the cell to expand, pressing against the cell wall. View larger image
Plants take up water from sources with a water potential higher than their own. For aquatic plants, the source is the surrounding aqueous medium. In freshwater environments, the presence of solutes in the plant's cells creates a water potential gradient from the surrounding water to the plant. In marine environments, plants must lower their water potential below that of seawater to take up water. Marine plants, as well as terrestrial plants of salt marshes and saline soils, adjust their osmotic potential in a manner similar to that of microorganisms by synthesizing solutes and taking up inorganic salts from their environment. Inorganic salts must be taken up selectively, however, because some, such as sodium (Na+) and chloride (Cl ), can be toxic at high concentrations.
The cell membranes of plants act as a solute filter, determining the amounts and types of solutes that move into and out of the plant.Terrestrial plants acquire water from the soil through their roots, as well as through associations with mutualistic fungi, called mycorrhizae, that grow into their roots from the soil (see Concept 15.1). The earliest land plants, which had not yet evolved roots, used mycorrhizal fungi to take up water and nutrients from the soil. The majority of modern terrestrial plant species use a combination of roots and mycorrhizae to take up water. Only the finest roots can take up water from the soil, because older, thicker roots develop a water-resistant waxy coating that limits their ability to absorb water as well as to lose water to the soil. Mycorrhizae provide a greater surface area for absorption of water and nutrients for the plant and allow greater exploration of the soil for these resources. In turn, the mycorrhizal fungi obtain energy from the plant.
Plants lose water by transpiration when their stomates open to allow carbon dioxide (CO2) from the atmosphere to diffuse into their leaves. Water moves out through the stomates, following the water potential gradient from the inside of the leaf (100% relative humidity) to the air. As we saw in the previous section, transpiration is an important cooling mechanism for leaves. The plant must replace the water lost by transpiration, however, if it is to avoid water stress. As a leaf loses water, the water potential of its cells decreases, creating a water potential gradient between the leaf and the xylem in the stem to which it is attached, so water moves through the xylem into the leaf. In this way, when the plant is transpiring, it creates a gradient of decreasing water potential from the soil through the roots and stems to the leaves (FIGURE 4.20). Water therefore flows from the soil, which has the highest water potential, into the roots, the xylem, and eventually the leaves, from which it is lost to the atmosphere via transpiration.
Because there is greater resistance to the movement of water into the roots and through the xylem than out through the stomates, the water supply from the soil cannot keep up with water loss by transpiration. As a result, the water content of the plant decreases during the day. At night the stomates close, and the water supply from the soil rehydrates the plant until it reaches near equilibrium with the soil water potential. This daily cycle of daytime dehydration and nighttime rehydration can go on indefinitely if the supply of water in the soil is adequate. The availability of water decreases when precipitation is not sufficient to replace the water lost from the soil through transpiration and evaporation. The water content of a plant will then decrease, and its turgor pressure will decrease as its cells become dehydrated (FIGURE 4.21). To avoid reaching a detrimentally or even lethally low water content, the plant must restrict its transpirational water loss. If leaf cells become so dehydrated that turgor is lost, the stomates close. This level of water stress can harm the plant, causing impairment of physiological functions such as photosynthesis. Extremely dry conditions can cause loss of xylem function.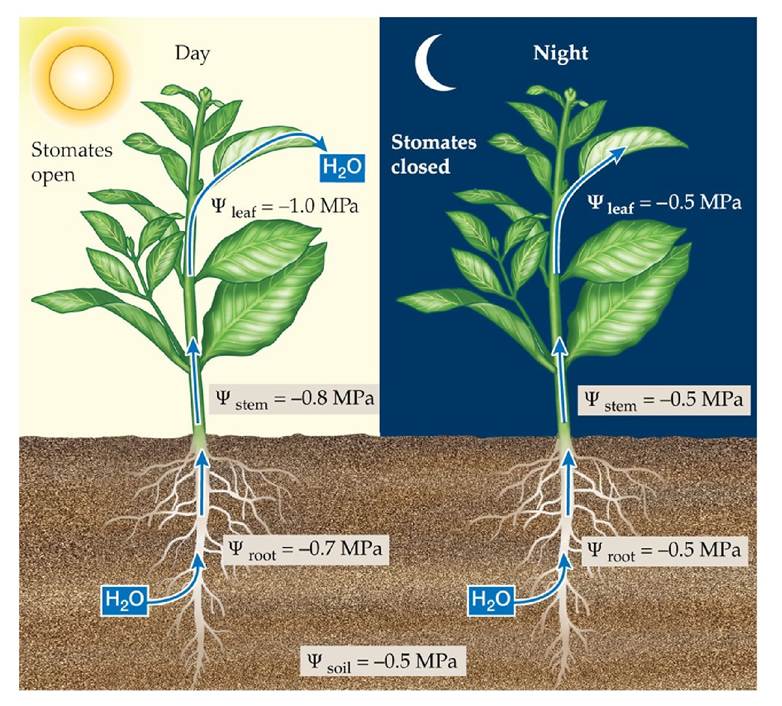
FIGURE 4.20 The Daily Cycle of Dehydration and Rehydration Duringtheday1Whenthe stomates are open, transpiration results in a gradient of water potential from leaf to stem, stem to roots, and roots to soil. At night, when the stomates are closed, water potential equilibrates as the plant rehydrates. View larger image
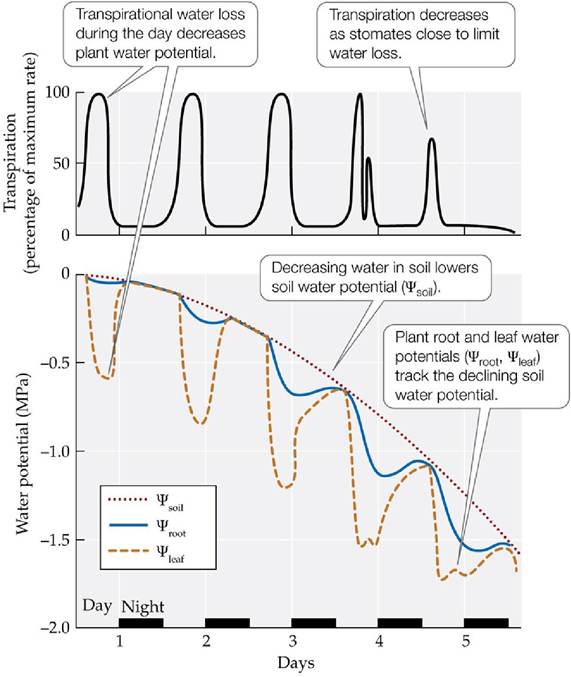
FIGURE 4.21 How Plants Cope with Depletion of Soil Water Ifsoilwaterisnot recharged, transpiration will deplete it, leading to progressive drying of the soil and a decrease in soil water potential.
As the soil dries, stomates may close at midday and reopen later in the afternoon, as seen on day 4 in the graph.
Assuming the air temperature is cooler later in the day, what influence would this have on plant water loss? (Top, after A. H. Fitter and R. K. Hay. 1987. Environmental Physiology of Plants. Academic Press: London; bottom, after R. D. Slatyer. 1967. Plant-Water Relationships. Academic Press: Cambridge, MA.) View larger imageSome plants of seasonally dry environments shed their leaves during long dry periods to eliminate transpirational water loss. Others have a signaling system that helps prevent the onset of water stress. As the soil dries out, the roots send a hormonal signal (abscisic acid) to the guard cells, which close the stomates, lowering the rate of water loss. Plants of dry environments, such as deserts, grasslands, and Mediterranean-type ecosystems, generally have better control of stomatal opening than plants of wetter climates. Plants of dry environments also have a thick waxy coating (cuticle) on their leaves to prevent water loss through the nonporous regions of the leaves. Additionally, plants of dry environments maintain a higher ratio of root biomass to biomass of stems and leaves than plants of moister environments, enhancing the rate of water supply to transpiring tissues (Mokany et al. 2006) (FIGURE 4.22). Some plants are capable of acclimatization by altering the growth of their roots to match the availability of soil moisture and nutrients.
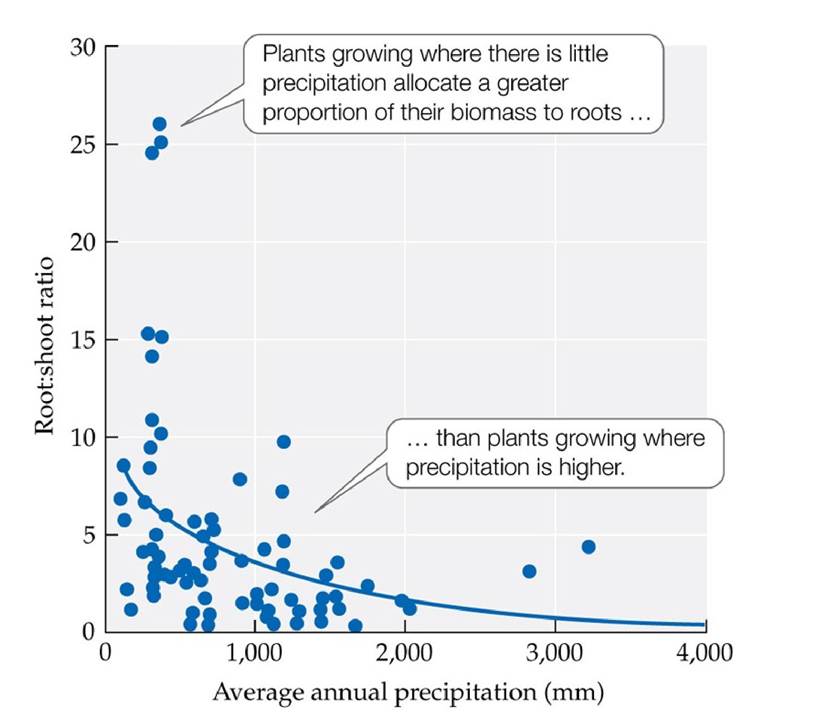
FIGURE 4.22 Allocation of Growth to Roots versus Shoots Is Associated with
Precipitation Levels The ratio of root biomass to leaf and stem (shoot) biomass increases with decreasing precipitation in shrubland and grassland biomes. Allocation of more biomass to roots in dry soils provides more water uptake capacity to support leaf function. (After K. Mokany et al.
2006. Global Change Biol 12: 84-96.) View larger image
Can plants have too much water? Technically, no, but saturation inhibits the diffusion of oxygen and can cause hypoxia in plant roots. Thus, waterlogged soils inhibit aerobic respiration in roots. Wet soils also enhance the growth of harmful fungal species that can damage roots. Ironically, the combination of these factors can lead to root death, which cuts off the supply of water to plants, and eventually to wilting. Adaptations to low oxygen concentrations in wet soils include root tissue containing air channels (called aerenchyma) as well as specialized roots that extend vertically above the water or waterlogged soil (as in mangroves; see Figure 3.19).
Water Balance in Animals
Multicellular animals face the same challenges plants and microorganisms do in maintaining water balance. Water losses and gains in animals, however, are governed by a more diverse set of exchanges than in plants and microorganisms (FIGURE 4.23). Many animals have the added complexity of specialized organs for gas exchange, ingestion and digestion, excretion, and circulation, all of which create areas of localized water and solute exchange as well as gradients of water and solutes within the animal's body. Most animals are mobile and can seek out environments conducive to maintaining a favorable water and solute balance, an option not available to plants or to most microorganisms.
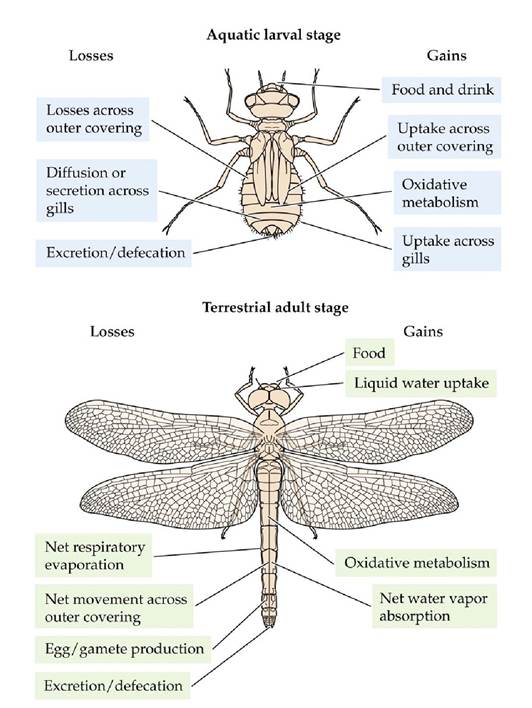
FIGURE 4.23 Gains and Losses of Water and Solutes in Aquatic and Terrestrial Animals Exemplified by Different Life Stages of a Dragonfly (After P. Willmer et al. 2005.
Environmental Physiology of Animals, 2nd ed. Blackwell Publishing: Malden, MA; E. B. Edney. 1980. In Insect Biology in the Future, M. Locke [Ed.], pp. 39-58. Academic Press: Cambridge, MA.) View larger image
Many animals must be able to maintain favorable water and solute balances under conditions of varying salinity. A marine animal that lacks this ability will die if transferred to brackish or fresh water. Although most marine invertebrates are isoosmotic to seawater, the specific types of solutes in their bodies can vary. Many invertebrates that are capable of adjusting to changes in the solute concentration of their environment do so by exchanging solutes with the surrounding seawater. Like plants, these animals must selectively control this exchange of specific solutes because some external solutes are toxic at the concentrations at which they are found in seawater and because some internal solutes are needed for biochemical reactions. Jellyfishes, squids, and crabs, for example, have Na+ and Cl- concentrations similar to those of seawater, but their sulfate (SO42-) concentrations may be one-half to one-fourth of those found in seawater.
Marine vertebrates include animals that are isoosmotic and hypoosmotic to seawater. The cartilaginous fishes, including the sharks and rays, have blood solute concentrations similar to those of seawater, although, as in invertebrates, their concentrations of specific solutes differ from those in seawater. In contrast, marine teleost (bony) fishes and mammals evolved in fresh water and later moved into marine environments. Their blood is hypoosmotic to seawater. Fish exchange water and salts with their environment through drinking and eating, and across the gills, which are also the organs of oxygen (O2) and CO2 exchange (FIGURE 4.24A). Salts that diffuse into or are ingested by marine teleost fishes must be continuously excreted in urine and through the gills against an osmotic gradient, which requires an expenditure of energy. Water lost across the gills must be replaced by drinking. Marine mammals, such as whales and porpoises, produce urine that is hyperosmotic to seawater and avoid drinking seawater to minimize salt uptake.
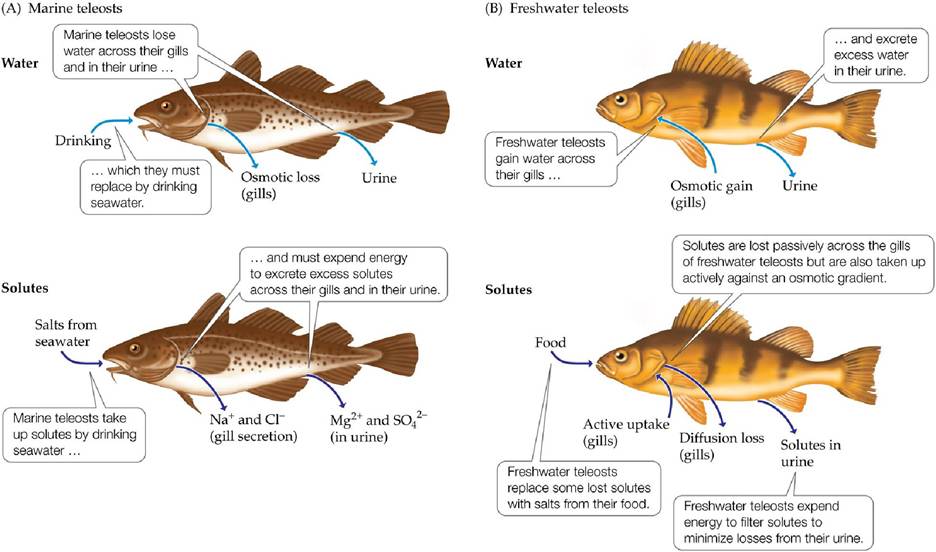
FIGURE 4.24 Water and Salt Balance in Marine and Freshwater Teleost Fishes Marine and freshwater teleost fishes face opposite challenges in maintaining water and solute balance.
(A) Marine teleosts are hypoosmotic to their environment: they tend to lose water and gain solutes. (B) Freshwater teleosts are hyperosmotic to their environment: they tend to gain water and lose solutes. (After K. Schmidt-Nielsen. 1979. Animal Physiology: Adaptation and Environment. Cambridge University Press: Cambridge.) View larger image
Freshwater animals are hyperosmotic relative to their environment; therefore, they tend to gain water and lose salts. Most salt exchange occurs at the gas exchange surfaces, including the skin of some invertebrates (e.g., freshwater worms) and the gills of many vertebrates and invertebrates. These animals must compensate for salt losses by taking up solutes in their food, and some groups, such as teleost fishes, must take up solutes actively through the gills against an osmotic gradient (FIGURE 4.24B). Excess water is excreted as dilute urine, from which the excretory system actively removes solutes to minimize their loss.
Terrestrial animals face the challenge of exchanging gases (O2 and CO2) in a dry environment with a very low water potential. These animals lower their evaporative water loss and exposure to water stress by having skin with a high resistance to water loss or by living in environments where they can compensate for high water losses with high water intake. Both approaches involve risks and trade-offs, however. A high resistance to water loss may compromise the animal's ability to exchange gases with the atmosphere. Reliance on a steady water supply puts the animal at risk if the source of water fails (e.g., during a severe drought). Tolerance for water loss varies substantially among groups of terrestrial animals. Generally, invertebrates have a higher tolerance for water loss than vertebrates. Within the vertebrates, amphibians have a higher tolerance for, but lower resistance to, water loss than mammals and birds (TABLE 4.1).
TABLE 4.1 Ranges of Tolerances for Water Loss in Selected Animal Groups
| Group | Weight loss (%) |
| Invertebrates | |
| Mollusks | 35-80 |
| Crabs | 15-18 |
| Insects | 25-75 |
| Vertebrates | |
| Frogs | 28-48 |
| Small birds | 4-8 |
| Rodents | 12-15 |
| Human | 10-12 |
| Camel | 30 |
Source: P. Willmer et al. 2005. Environmental Physiology of Animals, 2nd ed. Blackwell Publishing: Malden, MA.
Note: Values are maximum percentages of body weight lost as water that can be tolerated, based on observations of a range of exemplary species in each group.
Amphibians, including frogs, toads, and salamanders, rely primarily on stable water supplies to maintain their water balance. They can be found in a wide variety of biomes, from tropical rainforests to deserts, as long as there is a reliable source of water, such as regular rains or ponds. Amphibians depend on gas exchange through the skin to a greater degree than other terrestrial vertebrates. Therefore, amphibian skin is often thin, with a low resistance to water loss (FIGURE 4.25). However, some adult amphibian species have adapted to dry environments by developing specialized skin with higher resistance to water loss. For example, the southern foam-nest tree frog (Chiromantis xerampelina), which occurs throughout Africa, has skin that resists water loss in a manner similar to that of lizards. To compensate for reduced gas exchange through the skin, it has a higher breathing rate (Stinner and Shoemaker 1987). As a group, tree frogs have higher skin resistance to water loss than ground frogs, reflecting their drier habitat. Some ground frogs of seasonally dry environments, such as the northern snapping frog (Cyclorana australis) of Australia, lower their rates of water loss by forming a “cocoon” of mucous secretions consisting of proteins and fats that increases resistance to water loss.
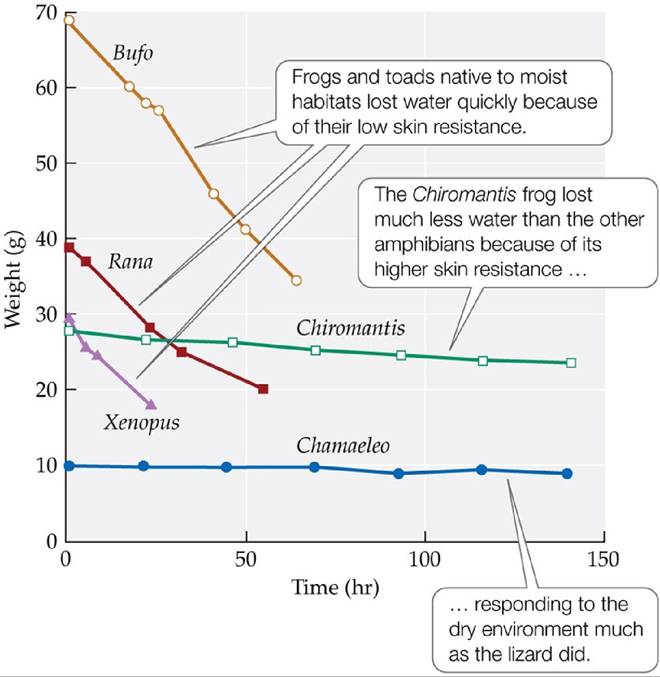
FIGURE 4.25 Resistance to Water Loss Varies among Frogs and Toads Amphibians were kept under uniform dry environmental conditions (25°C, 20%-30% relative humidity) to examine their rates of water loss, measured as loss of body weight. A lizard (Chamaeleo) was also tested for comparative purposes.
How could you estimate the resistances of these species to water loss quantitatively using this graph?
(After K. Schmidt-Nielsen. 1979. Animal Physiology: Adaptation and Environment. Cambridge University Press: Cambridge; based on J. P. Loveridge. 1970. Arnoldia [Rhodesia] 5: 1-6. National Museum of Southern Rhodesia.) View larger image
Reptiles have been extremely successful at inhabiting dry environments. The thick skin of desert snakes and lizards provides protection for the internal organs as well as an effective barrier to water loss. The outer skin, made up of multiple layers of dead cells with a fatty coating, is overlain by plates or scales. These layers give reptilian skin a very high resistance to water loss. Mammals and birds have skin anatomy similar to that of reptiles but have hair or feathers covering the skin rather than scales. The presence of sweat glands in mammals represents a trade-off between resistance to water loss and evaporative cooling. The highest resistances to water loss among terrestrial animals are found in the arthropods (e.g., insects and spiders), which are characterized by an outer exoskeleton made of hard chitin and coated with waxy hydrocarbons that prevents water movement (TABLE 4.2).
TABLE 4.2 Ranges of Resistance of External Coverings (Skin, Cuticle) to Water Loss
| Group | Resistance (s/cm) |
| Crabs (marine) | 6-14 |
| Fish | 2-35 |
| Frogs | 3-100 |
| Earthworms | 9 |
| Birds | 50-158 |
| Desert tortoises | 120 |
| Desert lizards | 1,400 |
| Desert scorpions, spiders | 1,300-4,000 |
Source: P. Willmer et al. 2005. Environmental Physiology of Animals, 2nd ed. Blackwell Publishing: Malden, MA.
An instructive example of how animals use a variety of integrated adaptations to cope with arid environments involves kangaroo rats (Dipodomys spp.), found throughout the deserts of North America. A combination of efficient water use and low rates of water loss greatly diminishes these rodents' water requirements (Schmidt-Nielsen and Schmidt-Nielsen 1951) (FIGURE 4.26). Kangaroo rats rarely drink water. A large proportion of their water requirement is met by eating dry seeds and by oxidative metabolism—that is, by metabolically converting carbohydrates and fats into water and carbon dioxide (Schmidt-Nielsen 1964). The animals also consume water-rich foods, such as insects or succulent vegetation, if they are available.
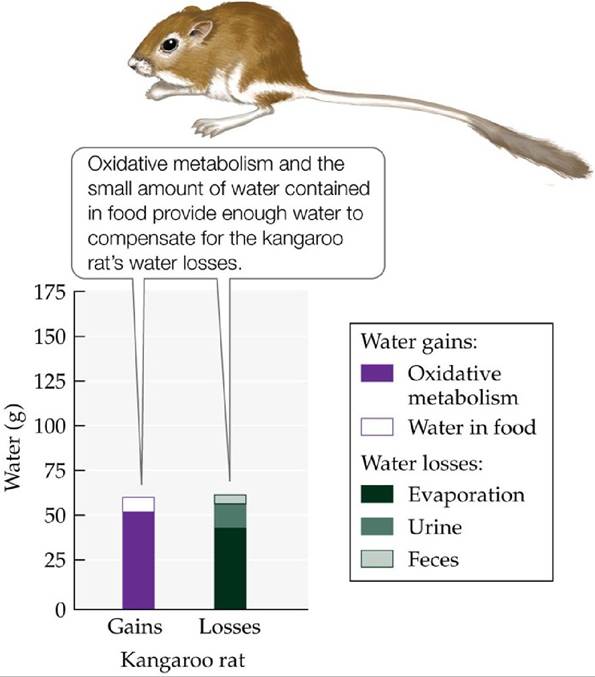
FIGURE 4.26 Water Balance in the Kangaroo Rat Under dry laboratory conditions (25°C, 25% relative humidity), kangaroo rats, native to deserts of western North America, do not require liquid water to survive. (After K. Schmidt-Nelson. 1997. Desert Animals. Clarendon Press: Oxford.) View larger image
Kangaroo rats minimize water loss through several physiological and behavioral adaptations. During the hottest periods of the year, they are active only at night, when air temperatures are lowest and humidities highest. During the day, they stay in their underground burrows, which are cooler and more humid than the desert surface. In some parts of their range, however, temperatures even in their burrows can rise high enough to expose kangaroo rats to significant evaporative water loss (Tracy and Walsberg 2002). To increase their resistance to this loss, kangaroo rats have thicker, oilier skin, with fewer sweat glands, than related rodents of moister environments. They minimize water losses in their urine and feces through effective removal of water by their kidneys and intestines. Kangaroo rats produce some of the most concentrated urine of any animal. The combination of these characteristics allows kangaroo rats to inhabit very arid environments without exposure to water stress, even without access to drinking water.
A Case Study Revisited
Frozen Frogs
The existence of amphibians above the Arctic Circle seems improbable, given their reliance on a steady supply of liquid water to maintain their water balance and the high potential for damage associated with freezing. Several problems must be overcome in order for complex organisms to survive freezing. First, when water freezes, it forms needlelike crystals that can penetrate and damage or destroy cell membranes and organelles. Second, the supply of oxygen to tissues is severely restricted by the lack of circulation and breathing. Finally, as ice forms, pure water is pulled from cells, resulting in shrinkage and an increase in solute concentration. Any one of these factors, or all of them working in combination, will kill tissues and organisms in subfreezing temperatures. Yet the frogs described in the Case Study, as well as many species of invertebrates, can tolerate the freezing of a substantial amount of their body water.
Wood frogs and other freeze-tolerant amphibians spend winter in shallow depressions under leaves, moss, or logs, which do not protect them from subfreezing temperatures. Several adaptations facilitate the survival of these amphibians through the winter and allow them to emerge from their frozen state in spring unharmed. Freezing of water in these animals is limited to the spaces outside the cells. A substantial proportion of their body water, from 35% to 65% in “fully frozen” frogs, freezes (Pinder et al. 1992). If more than 65% of their body water is frozen, most individuals will die because of excessive cell shrinkage. The formation of ice outside the cells is enhanced by the existence of ice-nucleating proteins that serve as the site of slow, controlled ice formation (Storey 1990). Solute concentrations in the unfrozen cells increase as the cells lose water to extracellular ice formation. In addition, freeze-tolerant amphibians synthesize additional solutes, including glucose and glycerol derived from the breakdown of liver glycogen. The resulting increase in solute concentrations lowers the freezing point inside the cells, allowing the intracellular solution to remain liquid at subfreezing temperatures. The concentrated solutes also stabilize the cell volume and the structures of organelles, proteins, and enzymes. As freezing proceeds, the frog's heart stops, and its lungs cease to pump air. Once it reaches this semistable state of partial freezing, the frog can remain frozen for several weeks, as long as the temperature does not drop below about -5°C (23°F). Although their winter “quarters” are not far below the surface of the ground, the insulating cover of leaves and snow keeps the frogs above that temperature.
The freezing process is initiated in wood frogs within minutes of ice formation within the animal, although the full process occurs over several days to weeks (Layne and Lee 1995). Thawing, on the other hand, may be rapid, with normal body functioning returning within 10 hours. This amazing amphibian feat of spending winter in a semifrozen state and emerging unharmed in spring has provided information to medical science that has facilitated the preservation of human tissues and organs at low temperatures (Costanzo et al. 1995), as well as optimism to proponents of whole-body cryonics, who hope that someday Grandpa can finally leave the Tuff Shed.
Connections in Nature
Desiccation Tolerance, Body Size, and Rarity
As we saw in Chapter 3, there is a close association between organisms’ adaptations to climate conditions and their distribution among terrestrial biomes. While subfreezing temperatures are an important constraint on the distribution and functioning of organisms in high-latitude and high-elevation biomes, low water availability is a more widespread challenge. Arid conditions can occur in most terrestrial biomes (see the climate diagrams in Concept 3.1), and they regularly occur over more than 60% of the land surface. As we have seen, the majority of terrestrial organisms, particularly animals, avoid exposure to dry conditions and rely on minimizing water losses to the environment. Some organisms, however, can tolerate arid conditions in much the same way that frozen frogs tolerate subfreezing winter conditions: by entering a dormant state while allowing themselves to dry out. This adaptive approach is common in microorganisms, including bacteria, fungi, and protists, but is also found in some multicellular animals and some plants, including mosses, liverworts, and a few flowering plants (Alpert 2006).
Desiccation-tolerant organisms can survive extreme dehydration, losing 80%-90% of their water as they equilibrate with the humidity of the air, then regain metabolic function shortly after they are rehydrated (FIGURE 4.27). As its cells dry out, the organism synthesizes sugars, which are the key to protecting its cell and organelle structures (Alpert 2006). Once dehydration proceeds beyond a certain threshold, metabolism ceases, and the sugars and the small amount of remaining water form a glassy coating over the cellular constituents. As with recovery from freezing, recovery from dehydration is rapid, occurring in hours to days.

FIGURE 4.27 Desiccation-TolerantOrganisms (A) The leaves of the club moss Selaginella Iepidophylla reach a very low moisture content during prolonged periods without rain (left); within 6 hours of receiving water, the leaves are functional and carrying out photosynthesis (right). (B) Water bears (tardigrades) are small invertebrates (less than 1 mm in length) found in aqueous environments, including oceans, lakes and ponds, soil water, and the water films on vegetation. Water bears contract and cease metabolism when they and their environment dry up (left) but rehydrate when moisture returns (right). View larger image
The prevalence of dry conditions in terrestrial environments suggests that desiccation tolerance should be more common than it is. Why hasn't such tolerance evolved in more plants and animals? A clue to this puzzle may be the small size of the organisms that are desiccation tolerant (Alpert 2006). Small organisms (less than 5 mm in animals) do not require structural reinforcements, such as a skeletal system, that would restrict the necessary shrinking of the organism as it dehydrates. In addition, water loss during dehydration must be slow enough to allow sugar synthesis to occur, but not so slow that the organism spends a long time with a low water content while metabolism is still occurring, which can cause physiological stress. Small organisms have surface-area-to-volume ratios and thicknesses favorable for the water loss rates required.
These arguments explain why desiccation tolerance is more common in small organisms, but not why they are rare (see Chapter 23). The two characteristics—small size and rarity—are intimately linked. As we will see in Chapter 14, small size is often associated with slow growth rates and poor competitive ability under conditions of low resource availability. Thus, natural selection for desiccation tolerance may involve trade-offs with other ecological characteristics, such as competitive ability, that might prevent these organisms from being successful in competitive environments.
2
- Many physiologists prefer using conductance rather than resistance to express the influence of a barrier on the movement of water or gases between an organism and its environment. Mathematically, conductance is the reciprocal of resistance.
More on the topic Water losses and solute gains and losses must be compensated:
- Previous versions of this work parted from joint losses distributions, considering market and credit losses.
- Equity implies a need for fairness in the distribution of gains and losses, and the entitlement of everyone to an acceptable quality and standard of living.
- CREDIT LOSSES
- SIMULATION OF JOINT LOSSES FOR THE BANKING SYSTEM
- SYSTEMIC RISK EXPOSURES AND LOSSES
- Economic Losses Associated with BTB in Nigeria
- Population Losses, 1926-1939
- Catchment studies measure losses of nutrients from ecosystems
- War Losses
- Estimating the Losses
- The System’s Distribution of Losses
- Nutrient Cycles and Losses
- Understanding the causes of diversity losses is a first step toward reversing them.
- Net primary production is the energy remaining after respiratory losses
- The field of conservation biology arose in response to global biodiversity losses
- Total loss-absorbing capacity (TLAC) considers the scope for a bank to absorb losses.