NUTRITIONAL, METABOLIC, AND TOXIC DISORDERS
Hypovitaminosis C: Scurvy
Deficiency of dietary vitamin C is a significant clinical entity in guinea pigs that may manifest itself in a number of ways. Ascorbic acid-dependent species are genetically deficient in the enzyme L-gulonolactone oxidase, which is involved in the conversion of L-gulonolactone to L-ascorbic acid.
This biosynthetic activity occurs in the liver in mammals, but the synthesis of vitamin C occurs in the kidney in amphibians and reptiles. In addition to the inability of simian and human primates and guinea pigs to synthesize endogenous vitamin C, certain bats (e.g., Indian fruit bat), some birds (e.g., red-vented bulbul bird, northern shrike), some fish (e.g., channel catfish), and cetaceans also require dietary sources of vitamin C. Ascorbic acid is essential in the hydroxylase reactions necessary for the formation of hydroxyproline and hydroxylysine in the collagen molecule. Thus, connective tissue cells are unable to synthesize collagen at a normal rate, resulting in deficient and defective production of interstitial osseous matrix. Vitamin C is also necessary for the catabolism of cholesterol to bile acids. In scurvy, cartilage produced in the physeal plates persists and lengthens, but it is not replaced by bone. This calcified cartilage scaffolding is relatively susceptible to mechanical forces; thus multiple microfractures occur in the epiphyseal region. Immobilization of the limb in a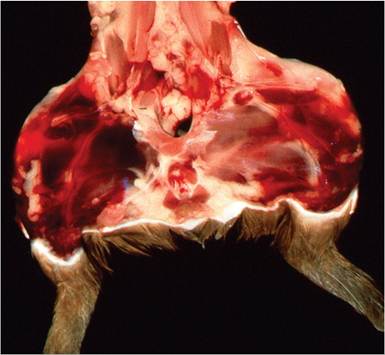
FIG. 5.32. Hypovitaminosis C (scurvy) in a young guinea pig. Note the periarticular hemorrhages in the region of the knees.
plaster cast will prevent the occurrence of microfractures, emphasizing the effect of the normal stresses and strains of limb movement on the development of lesions. There is increased capillary fragility, with widening of intercellular spaces between endothelial cells, vacuolar degeneration of endothelium, and depletion of subendothelial collagenous tissue.
There is also increased prothrombin time in animals with scurvy. The increased susceptibility of scorbutic guinea pigs to bacterial infections such as S. pneumoniae is probably due, at least in part, to impaired macrophage migration and depressed phagocytic activity of heterophils.Pathology
Hemorrhages are present in the periarticular regions, particularly in the hind limbs (Fig. 5.32), but may also appear randomly in other tissues. There may be enlargement of the costochondral junctions (scorbutic rosary, Fig. 5.33), with hemorrhages into the regional soft tissues. Animals may be thin and appear unkempt. Evidence of diarrhea is a variable finding. Occasionally,
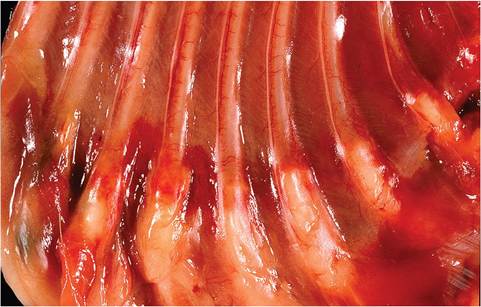
FIG. 5.33. Enlarged costochondral junctions (scorbutic rosary) and regional hemorrhages in a young guinea pig with hypovitaminosis C.
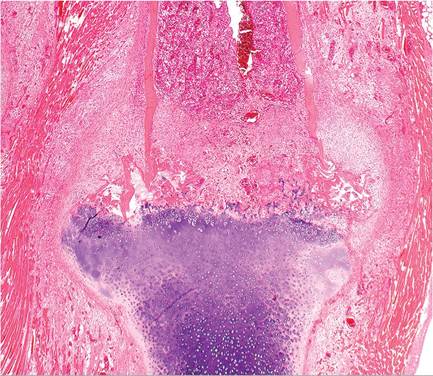
FIG. 5.34. Costochondral junction from a case of scurvy in a guinea pig. Note the callus formation of the junctional periostieum, persistence of cartilage with failure of ossification, microfractures of bone, and marked proliferation of fusiform mesenchymal cells in the periosteal and medullary regions.
blood-tinged gut contents are observed, and there may be ecchymoses in the urinary bladder. Adrenal glands are frequently markedly enlarged. Microscopically, persistence and irregularities of the physeal cartilage is evident in young growing animals. Microfractures of the cartilaginous spicules and hemorrhage are common findings. In bones including ribs, there is marked proliferation of poorly differentiated fusiform mesenchymal cells in the periosteal regions (Figs. 5.34 and 5.35) and medullary cavity, with displacement of normal hematopoietic cells. Frequently, there are aggregations of eosinophilic material interspersed between the mesenchymal cells. Dental abnormalities also occur.
Fibrosis of the pulp and derangement of odontoblasts have been observed during the early stages of the disease. Hemosiderin-laden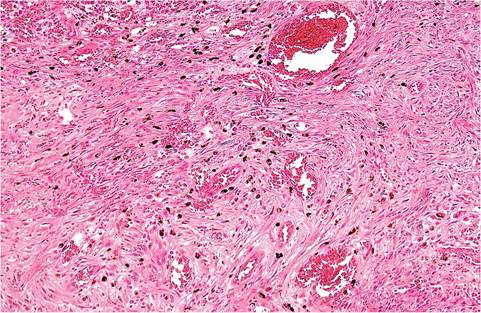
FIG. 5.35. Higher magnification of mesenchymal proliferation from
the case depicted in the previous figure. Note the hemorrhage and numerous hemosiderin-laden cells throughout the proliferating mesenchyma.
macrophages that are commonly present in the lamina propria of the intestine have been considered by some to be due to subclinical hypovitaminosis C.
Myopathies Nutritional Myopathy
Guinea pigs are prone to development of degenerative and necrotizing myopathy. The etiology may not be determined in many cases. However, there is a clear relationship to selenium/vitamin E deficiency. Myopathy can be readily induced in guinea pigs with selenium/ vitamin E deficient diets, and natural cases of myopathy have been reversed by dietary change and treatment with selenium and alphatocopherol. Experimental studies have shown that combined selenium and vitamin E deficiency causes a fatal myopathy that is more severe than the myopathy that arises with vitamin E alone. Furthermore, combined selenium and vitamin C deficiency causes more severe myopathy than selenium alone.
Depression and conjunctivitis may be present on clinical examination. In 1 report, spontaneous hind limb weakness was a prominent clinical feature of the disease. Severely affected animals may die within 1 week of the onset of clinical signs. Elevated serum creatine phosphokinase (CPK) is a feature of the disease. At necropsy, there is a marked pallor of the affected muscles. Microscopic changes are characterized by coagulative necrosis and hyalinization of cardiac and skeletal myofibers, fragmentation of sarcoplasm, increased basophilia of the sarcoplasm, and rowing of nuclei in regenerating myofibers (Fig. 5.36). Multinucleated muscle fibers may be present in regenerating myofibers.
Mineralization of myofibers is apparently not an important feature of the disease. There may be marked reduction in reproductive performance in affected sows, and fetuses of vitamin E-deficient sows may develop encephaloma- lacia. Testicular degeneration is a later development seen in vitamin E-deficient boars.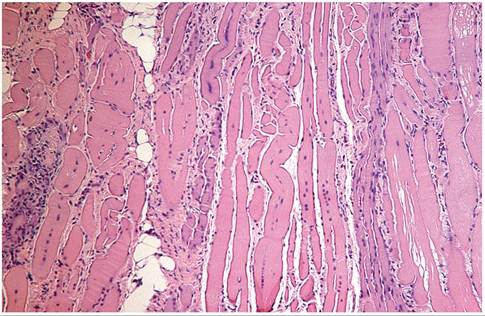
FIG. 5.36. Myocardium from an adult guinea pig with multiphasic degeneration and regeneration of myofibers.
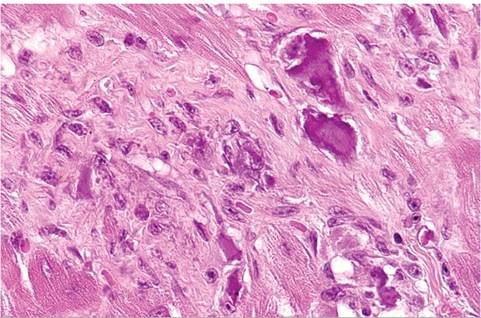
FIG. 5.37. Focal myocardial degeneration and fibrosis with mineralization in an aged guinea pig.
Myocardial and Skeletal Muscle Degeneration with Mineralization
This is a poorly understood syndrome, and the contributing factors have not been clearly identified. Multifocal mineralization/calcification of individual muscle fibers may be seen as an incidental finding, particularly in the major muscles of the hind limbs. Affected animals frequently are subclinical. On microscopic examination, there may be multifocal mineralization of skeletal myofibers, and less commonly cardiac myofibers. Changes are characterized by degeneration of myofibers, with variable mineralization and minimal mononuclear cell infiltration. In chronic lesions of longer duration, there may be concurrent mineralization with fibrosis (Fig. 5.37). In 1 report, myocardial lesions were observed in crossbred Abyssinian/Hartley guinea pigs. Vitamin E and selenium levels were within normal limits, and genetic factors were implicated in the disease.
Cardiac Glycogenosis: Rhabdomyomatosis
This condition is observed as an incidental finding in guinea pigs of various ages. It has been interpreted to be a degenerative condition and a congenital tissue malformation with "blastemoid" characteristics. It has been speculated but not proven that rhabdomyomatosis occurs more frequently in animals with scurvy.
The current assessment is that rhabdomyomatosis is related to a disorder of glycogen metabolism.Smaller lesions are not visible on macroscopic examination. Occasionally, larger areas appear as pale pink, poorly delineated foci or streaks. These changes have been observed in various regions of the heart, including ventricles, atria, interventricular septa, and papillary muscles. Lesions are most frequently found in the left ventricle. Microscopic examination reveals a spongy network of vacuolated myofibers composed of finely fibrillar to granular, eosinophilic cytoplasm (Fig. 5.38). Vacuoles are rounded to polygonal in shape and usually fill the sarcolemmal sheath. Vacuoles contain large quantities of glycogen, which is washed out in the
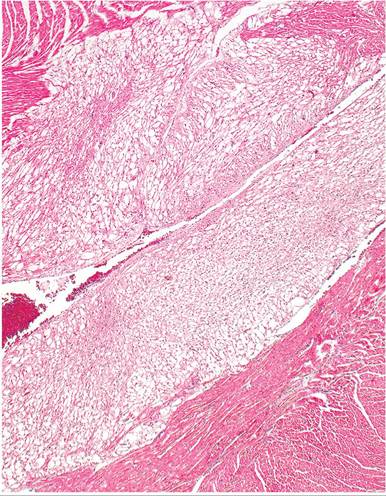
FIG. 5.38. Heart from an adult guinea pig with focal myocardial rhabdomyomatosis, in which foci of myofibers are pale staining due to distention of sarcoplasm with glycogen, which is lost during histologic processing.
fixation and processing procedures (Fig. 5.39). Glycogen is readily demonstrated in PAS-stained, alcohol-fixed specimens. There may be displacement and flattening of myocyte nuclei in some affected fibers. In other fibers, there may be a cytoplasmic marginal rim with a round nucleus projecting into the vacuole. Myofibers with centrally located nuclei and radiating fibrillar processes have been called “spider cells.” Interspersed within the affected myofibers, there may be poorly differentiated fibers with identifiable cross-striations.
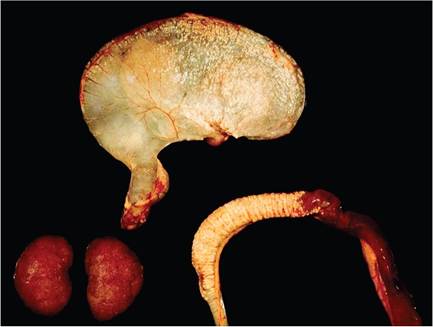
FIG. 5.40. Metastatic mineralization of the gastric and intestinal walls in an aged guinea pig with chronic kidney disease (lower left). There are chalky deposits on the serosal surface of the stomach and intestine.
may be confined to soft tissues around the elbows and ribs. There may be more widespread mineralization of tissues, such as lung, trachea, heart, aorta, liver, kidney, stomach, uterus, and sclera (Figs.
5.40 and 5.41). Dietary factors such as low magnesium and high phosphorus have been implicated in this syndrome. High-calcium or high-phosphorus diets appear to interfere with magnesium absorption and metabolism. Therefore, this syndrome may not be the result of a deficiency of a single component but rather may be due to a dietary imbalance of 2 or more nutrients. As with the rabbit, renal excretion of calcium is an important regulator of serum calcium levels, so metastatic mineralization may be associated with renal disease.Metastatic Mineralization
Metastatic mineralization occurs most often in guinea pigs over 1 year of age. Muscle stiffness and unthriftiness are variable findings. In some cases, mineral deposition
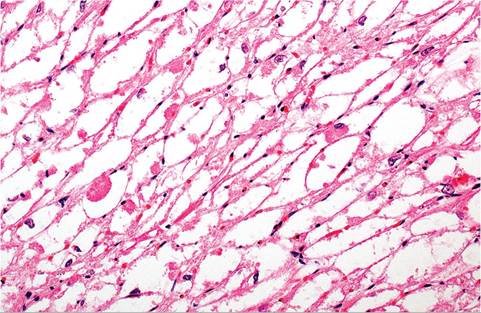
FIG. 5.39. Higher magnification of affected myocardium with rhabdomyomatosis.
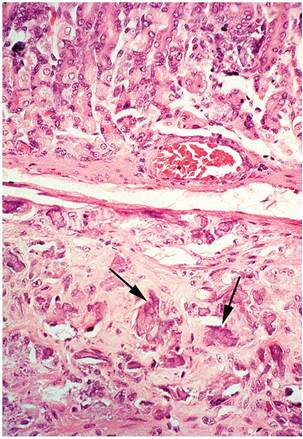
FIG. 5.41. Gastric wall with metastatic mineralization of the smooth muscle (arrows).
Pregnancy Toxemia
Although the clinical signs are similar in many respects, there are 2 different patterns of disease associated with pregnancy toxemia in the guinea pig: the fasting or metabolic form and the circulatory or toxic form. Both forms normally occur in advanced pregnancy. Depression, acidosis, ketosis, proteinuria, ketonuria, and a lowered urinary pH from around 9 (normal) to 5-6 are frequent manifestations in both forms of pregnancy toxemia.
Metabolic Form
This form of pregnancy toxemia occurs in obese sows during the last 2-3 weeks of pregnancy, particularly in sows during their first or second pregnancy. The uterine contents of guinea pigs in advanced pregnancy may represent up to 50% of the weight of the nonpregnant dam. Stress factors such as shipping or changes in feeding routines may be sufficient to precipitate the disease. In 1 study, withholding the usual supplemental cabbage ration resulted in a mortality rate of approximately 5% in obese dams. The syndrome has also been produced in obese, nonpregnant animals subjected to the stress of cabbage deprivation. Lowered blood glucose, ketosis, and hyperlipidemia are typical findings. Animals usually become comatose and die within 5-6 days after the onset of the disease. It appears to be triggered by high-energy feeding followed by subsequent withdrawal, resulting in mobilization of fat as a source of energy, with disastrous results. At necropsy, animals usually have abundant fat reserves, with marked hepatic lipidosis. Microscopically, marked lipidosis is evident in hepatocytes (Fig. 5.42), as well as in kidney and adrenals. Lipid may also be demonstrable in vessels with fat stains.
Circulatory/Toxic Form: Preeclampsia
In this form of pregnancy toxemia, uteroplacental ischemia may occur due to compression of the aorta caudal to the renal vessels by the gravid uterus. This results in a significant reduction in blood pressure in the uterine vessels, with subsequent placental necrosis and hemorrhage,
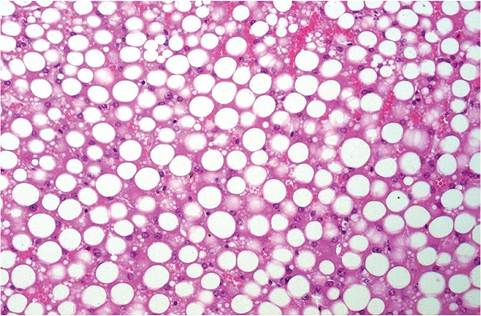
FIG. 5.42. Hepatic lipidosis in a sow with the metabolic form of pregnancy toxemia.
thrombocytopenia, ketosis, and death. On microscopic examination, there is uterine and placental hemorrhage, necrosis, and leukocytic infiltration. Multifocal periportal liver necrosis, nephrosis, and adrenocortical hemorrhage are typical findings. The disease has been reproduced in female guinea pigs by banding and transection of uterine and ovarian vessels.
Diabetes Mellitus
Spontaneous diabetes mellitus can arise in guinea pigs. Affected animals frequently show no clinical signs during the early stages of the disease. In 1 report, animals were usually affected by 6 months of age, and the average age of onset was 3 months. Both sexes were affected. Changes evident by clinical chemistry were hyperglycemia, glycosuria, and rarely ketonuria. There was a marked reduction in fertility in affected sows. Animals introduced into the affected colony subsequently became diabetic. An unidentified infectious agent was suspected to be involved. On microscopic examination, there was vacuolation of and degranulation of the beta islet cells with fatty vacuolation of exocrine cells and fibrosis of the vascular stroma. The exocrine component was found to result in reduced overall secretion, bicarbonate concentration, and enzyme production. In advanced cases, there was thickening of basement membranes of the glomerular tufts, sometimes with sclerosis and scarring of Bowman's capsule.
Systemic Amyloidosis
Amyloidosis is occasionally found in guinea pigs as an incidental finding at necropsy. It typically involves deposition of hyaline AA amyloid in the periphery of splenic follicles, between hepatic cords and sinusoids, kidney (Figs. 5.43 and 5.44), and adrenal cortex. Other organs
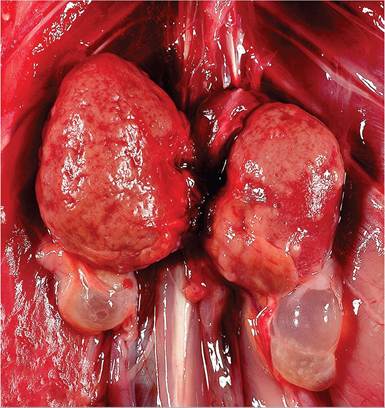
FIG. 5.43. Renal amyloidosis in an aged guinea pig. Also note the bilateral cystic rete ovarii below the kidneys. (Source: R. Burns, University of Connecticut, Mansfield, Connecticut. Reproduced with permission from R. Burns.)
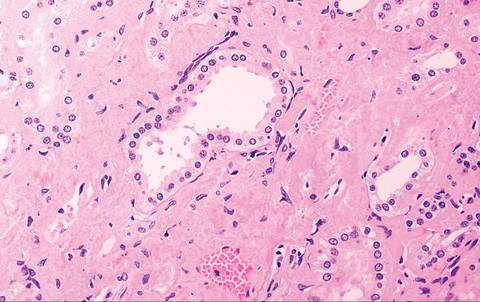
FIG. S.44. Amyloid deposition in the renal cortical Intersitium in the aged guinea pig depicted in the previous figure.
may be involved less commonly. It tends to occur in immunized guinea pigs and in animals with chronic bacterial infections, such as pododermatitis.
Toxic Disorders
Guinea pigs are subject to a number of ingested toxins, similar to rabbits. Their chewing behavior makes them prone to chronic lead poisoning from lead-containing environmental surfaces. A large number of poisonous plants pose a risk to guinea pigs, including philodendron and peace lily, which contain oxylates that result in renal failure.
BIBLIOGRAPHY FOR NUTRITIONAL, METABOLIC, AND TOXIC DISORDERS
General Reference
Williams, B. (2012) Non-infectious diseases. In: The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents (eds. M. A. Suckow, K. A. Stevens, & R. P. Wilson), pp. 685-704. Elsievier, London.
Hypovitaminosis C: Scurvy
Clarke, G.L., Allen, A.M. Small, J.D., & Lock, A. (1980) Subclinical scurvy in the guinea pig. Veterinary Pathology 17:40-44.
Eva, J.K., Fifield, R., & Rickett, M. (1976) Decomposition of supplementary vitamin C in diets compounded for laboratory animals. Laboratory Animals 10:157-159.
Follis, R.H. (1943) Effect of mechanical force on the skeletal lesions in acute scurvy in guinea pigs. Archives of Pathology 35:579-582.
Ganguly, R., Durieux, M.F., & Waldman, R.H. (1976) Macrophage function in vitamin C-deficient guinea pigs. American Journal of Clinical Nutrition 29:762-765.
Gillespie, D.S. (1980) An overview of species needing vitamin C. Journal of Zoo Animal Medicine 11:88-91.
Gore, I., Fujinami, T., & Shirahama, T. (1965) Endothelial changes produced by ascorbic acid deficiency in guinea pigs. Archives of Pathology 80:371-376.
Kim, J.C.S. (1977) Ultrastructural studies of vascular and muscular changes in ascorbic acid-deficient guinea pigs. Laboratory Animals 11:113-117.
Nungester, W.J. & Ames, A.M. 1948. The relationship between ascorbic acid and phagocytic activity. Journal of Infectious Diseases 83:50-54.
Myopathies
Griffith, J.W. & Lang, C.M. (1987) Vitamin E and selenium status of guinea pigs with myocardial necrosis. Laboratory Animal Science 37:776-779.
Hill, K.E., Motley, A.K., Li, X., May, J.M., & Burk, R.F. (2001) Combined selenium and vitamin E deficiency causes fatal myopathy in guinea pigs. Journal of Nutrition 131:1798-1802.
Hill, K.E., Motley, A.K., May, J.M., & Burk, R.F. (2009) Combined selenium and vitamin C deficiency causes cell death in guinea pig skeletal muscle. Nutritional Research 29:213-219.
Howell, J.M. & Buxton, P.H. (1975) Alpha tocopherol responsive muscular dystrophy in guinea pigs. Neuropathology and Applied Neurobiology 1:49-58.
Hueper, W.C. (1941) Rhabdomyomatosis of the heart in a guinea pig. American Journal of Pathology 17:121-126.
Pappenheimer, A.M. & Schogoleff, C. (1944) The testis in vitamin E deficiency in guinea pigs. American Journal of Pathology 20:239-244.
Saunders, L.Z. (1958) Myositis in guinea pigs. Journal of the National Cancer Institute 20:899-903.
Takahashi, M., Iwata, S., Matsuzawa, H., & Fujiwara, H. (1985) Pathological findings of cardiac rhabdomyomatosis in the guinea pig. Jikken Dobutsu 34:417-424.
Vink, H. (1969) Rhabdomyomatosis of the heart in guinea pigs. Journal of Pathology 97:331-334.
Ward, G.S., Johnsen, D.O., Kovatch, R.M., & Peace, T. (1977) Myopathy in guinea pigs. Journal of the American Veterinary Medical Association 171:837-838.
Webb, J.N. (1970) Naturally occurring myopathy in guinea pigs. Journal of Pathology 100:155-162.
Metastatic Mineralization
Galloway, J.H., Glover, D., & Fox, W.C. (1964) Relationship of diet and age to metastatic calcification in guinea pigs. Laboratory Animal Care 14:6-12.
Sparschu, G.L. & Christie, R.J. (1968) Metastatic calcification in a guinea pig colony: a pathological survey. Laboratory Animal Care 18:520-526.
Pregnancy Toxemia
Bergman, E.N. & Sellers, E.F. (1960) Comparison of fasting ketosis in pregnant and nonpregnant guinea pigs. American Journal of Physiology 198:1083-1086.
Ganaway, J.R. & Allen, A.M. (1971) Obesity predisposes to pregnancy toxemia (ketosis) of guinea pigs. Laboratory Animal Science 21:40-44.
Golden, J.G., Hughes, H.C., & Lang, C.M. (1980) Experimental toxemia in the pregnant guinea pig (Cavia porcellus). Laboratory Animal Science 30:174-179.
Lachmann, G., Hamel, I., Holdt, J., & Furll, M. (1989) The fat mobilization syndrome of guinea pigs (Caviaporcellus L.). Archiv fur Experimentelle Veterinarmedizin 43:231-240.
Seidl, D.C., Hughes, H.C., Bertolet, R., & Lang, C.M. (1979) True pregnancy toxemia (preeclampsia) in the guinea pig (Cavia porcellus). Laboratory Animal Science 29:472-478.
Diabetes Mellitus
Balk, M.W., Lang, C.M., White, W.J., & Munger, B.L. (1975) Exocrine pancreatic dysfunction in guinea pigs with diabetes melli- tus. Laboratory Investigation 32:28-32.
Lang, C.M., Munder, R.L., & Rapp, F. (1977) The guinea pig as a model of diabetes mellitus. Laboratory Animal Science 27:789-805.
Langner, P.H., Lang, C.M., Singh, S.B., Munger, B.L., & Abt, A.B. (1981) Glomerular basement membrane changes in aging nondiabetic and diabetic guinea pigs. Experimental Aging Research 7:93-105.
Munger, B.L. & Lang, C.M. (1973) Spontaneous diabetes mellitus in guinea pigs: the acute Cytopathology of the islets of Langerhans. Laboratory Investigation 29:685-702.
Systemic Amyloidosis
Pirani, C.L., Bly, C.G., Sutherland, K., & Chereso, F. (1949) Experimental amyloidosis in the guinea pig. Science 110:145-146.
Taylor, J.L., Wagner, J. E., Owens, D.R., & Shulman, R.A. (1971) Chronic pododermatitis in guinea pigs. Laboratory Animal Science 21:944-945.
Toxic Disorders
Gfeller, R.W. & Messonnier, S.P. (2004) Handbook of Small Animal Toxicology and Poisonings, 2nd edn. Mosby, St. Louis, MO.
Holowaychuk, M.K. (2006) Renal failure in a guinea pig (Cavia porcellus) following ingestion of oxalate containing plants. Canadian Veterinary Journal 47:787-789.
More on the topic NUTRITIONAL, METABOLIC, AND TOXIC DISORDERS:
- NUTRITIONAL, METABOLIC AND TOXIC DISORDERS
- NUTRITIONAL AND METABOLIC DISORDERS
- METABOLIC DISORDERS IN NEWBORN
- Nutritional Disorders
- Metabolic Disorders
- Chapter 6 METABOLIC DISORDERS
- GENETIC, METABOLIC, AND OTHER DISORDERS
- Antiretroviral Therapy and Metabolic Disorders
- A metabolic disease, or metabolic disorder, is the name given to a group of illnesses in dairy cows which are caused by an over-exertion of their normal metabolism.
- Toxic Alcohols
- Toxic Shock Syndrome
- Toxic Neuropathies
- TOXIC CAUSES