Trophic interactions can trickle down through multiple trophic levels
Changes in abundance or species composition at one trophic level can lead to important changes in abundance and species composition at other trophic levels. For example, an increase in the rate of predation by a carnivore at the fourth trophic level on carnivores at the third trophic level would lead to a lower rate of consumption of herbivores at the second trophic level.
More herbivory would decrease the abundance of autotrophs and would therefore lower rates of NPP. Nonconsumptive species interactions, such as competition, can have similar topdown effects on abundance and species composition at lower trophic levels, as we'll see shortly. Such a series of changes in abundance and species composition is referred to as a trophic cascade.Our understanding of trophic cascades comes primarily from aquatic ecosystems, although there are examples from terrestrial ecosystems as well. Several generalizations have been drawn from studies of these interactions. First, trophic cascades are most often associated with changes in the abundance of top specialist predators. Second, omnivory may act to buffer the effects of trophic cascades through the consumption of prey at multiple trophic levels. Finally, trophic cascades have been hypothesized to be most important in relatively simple, species-poor ecosystems. However, several recent experiments have demonstrated trophic cascades in ecosystems with relatively high species diversity.
An Aquatic Trophic Cascade
There are several studies of trophic cascades associated with near extinctions of native species. A classic example is the interaction among sea otters (Enhydra lutris), sea urchins, and killer whales on the west coast of North America, which was discussed in the Case Study Revisited in Chapter 9. Additionally there are numerous examples of trophic cascades associated with the intentional or unintentional introduction of non-native species.
One such example resulted from the release of brown trout (Salmo trutta), a popular sport fish, into streams and lakes of New Zealand. The stocking of Kiwi waters by European settlers began in the 1860s, and by 1920 an estimated 60 million fish had been released throughout New Zealand. Native fish populations have declined as a result, and some species have disappeared from streams now dominated by trout.Alexander Flecker and Colin Townsend (1994) investigated the influence of the brown trout on the species composition of its prey (primarily stream insects) and associated effects on primary production in the Shag River. Brown trout were originally released into the Shag River in 1869 by the “Otago Acclimatisation Society” to make settlers feel more at home. The Shag River is one of a small number of streams in New Zealand that still holds both native fish and trout in the same sections. Native fish species include the common river galaxias (Galaxias vulgaris). The morphology and feeding behavior of galaxias are similar to those of trout, as indicated by the common name for the galaxias, Maori trout.
Flecker and Townsend compared the effects of brown trout and galaxias on stream invertebrate species composition and abundance as well as on primary production by algae. To manipulate fish presence and absence, they constructed artificial stream channels adjacent to the natural channel, made of 5-m lengths of half-cylinders of PVC pipe. The PVC channels had mesh on the ends that kept fish in or out but allowed free movement of stream invertebrates and algae. The researchers placed clean gravel and stone cobbles in the bottoms of the channels to provide a substrate for the invertebrates and algae. The channels were allowed to accumulate algae and invertebrates for 10 days before the fish were added. Three treatments were initiated: channels with introduced brown trout, channels with galaxias, and channels with no fish (controls). Eight fish of similar size and mass were used for each fish species addition.
The experiment was run for 10 days, after which samples were collected to determine invertebrate species composition and abundance and algal biomass.Flecker and Townsend had expected brown trout to decrease invertebrate diversity more than the native galaxias, but the effect of fish on invertebrate diversity was relatively small and did not differ between the two fish species. The brown trout, however, reduced total invertebrate density by approximately 40% relative to the control channels, while galaxias resulted in a smaller reduction (FIGURE 21.10A). The abundance of algae increased with both fish, but the effect was greater in the channels with trout (FIGURE 21.10B). Flecker and Townsend suggested that the effect on algal biomass was the result of a trophic cascade in which fish predation not only reduced the density of stream invertebrates, but also caused them to spend more time in refugia on the stream bottom rather than feeding on algae. The trout had a much greater effect on invertebrate density, and thus on primary production, than the native galaxias. These results suggested that trophic cascades associated with the stocking of non-native fish for sport may have consequences not just for native biodiversity, but for the functioning of stream ecosystems as well.
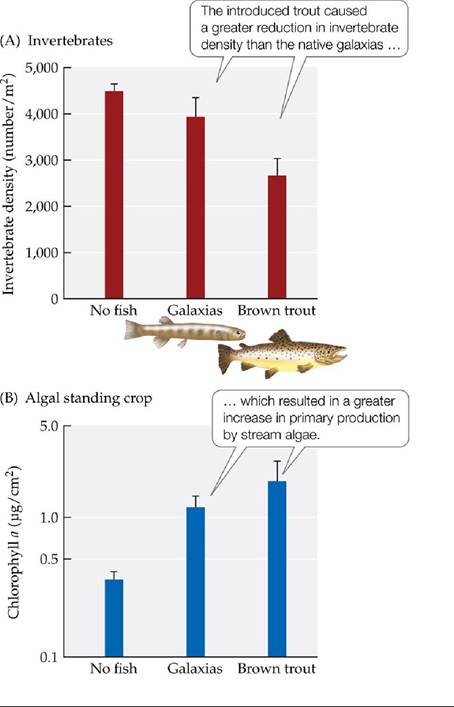
FIGURE 21.10 AnAquaticTrophicCascade FleckerandTownsendusedartificialstream channels to study the effects of non-native brown trout and a native fish (galaxias) on stream invertebrates and algae in the Shag River, New Zealand. (A) Effects on invertebrate density. (B) Effects on algal biomass, as estimated using chlorophyll concentrations in stream water. Error bars show one SE of the mean.
What factor other than overall consumption rate might explain why the presence of brown trout results in a larger increase in primary production than the presence of native galaxias?
(After A. S.
Flecker and C. R. Townsend. 1994. Ecol Appl 4: 798-807.) View larger imageA Terrestrial Trophic Cascade
As mentioned earlier, trophic cascades have been most commonly observed in aquatic ecosystems, where they are more frequent and their effects are stronger than in terrestrial ecosystems (Shurin et al. 2002). Terrestrial ecosystems are generally thought to be more complex than aquatic ecosystems. In addition, it was believed that a decrease in the abundance of one species in a terrestrial ecosystem was more likely to be compensated for by an increase in the abundances of similar species that were not being consumed as heavily. Thus, trophic cascades were considered unlikely in diverse terrestrial ecosystems such as tropical forests.
Lee Dyer and Deborah Letourneau (1999a) tested the effects of a potential trophic cascade on the production of Piper cenocladum trees in the understory of a lowland tropical rainforest in Costa Rica. Piper cenocladum is a relatively common component of the understory in these forests and is eaten by dozens of different herbivore species. Ants of the genus Pheidole live in chambers in the petioles of the leaves of the Piper trees. The ants eat food bodies provided by the trees, and they also consume herbivores that attack the trees. These ants, in turn, are eaten by beetles of the genus Tarsobaenus. Thus, four distinct trophic levels exist in this system (FIGURE 21.11). Dyer and Letourneau had previously noted that plant biomass was lower, and rates of herbivory were higher, when densities of Tarsobaenus beetles were high. They performed experiments to test whether a trophic cascade involving the beetles, ants, and herbivores influenced the production of the Piper trees and how strong that influence was, compared with that of bottom-up factors such as light and soil fertility.
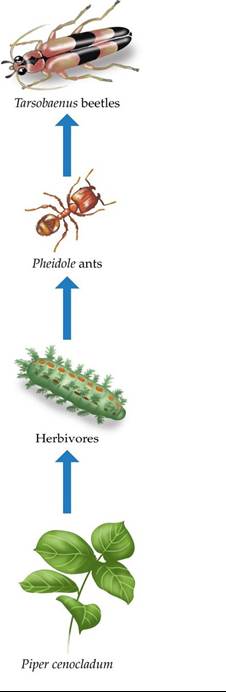
FIGURE 21.11 ATerrestrialTrophicCascade Trophicinteractionsintheunderstory ecosystem of a lowland tropical rainforest in Costa Rica.
Piper cenocladum trees are consumed by herbivores but provide shelter for Pheidole ants, which consume herbivores attacking the trees. Pheidole ants are consumed by Tarsobaenus beetles. Both ants and beetles also consume food bodies produced by the trees. (After L. A. Dyer and D. K. Letourneau. 1999a. Proc Natl Acad Sci USA 96: 5072-5076. © 1999 National Academy of Sciences, U.S.A.) View larger imageDyer and Letourneau established experimental plots in the understory by planting uniform-sized cuttings of Piper trees. They treated two groups of plots with an insecticide to kill any ants present, then added Tarsobaenus beetle larvae to one of those groups of plots. This procedure established three groups of treatment plots: one group of insecticide-treated plots with beetles, one control group of insecticide-treated plots without beetles, and one control group of untreated plots. In the plots with beetles, the insecticide treatment facilitated the establishment of the beetles by preventing ant attacks on the beetle larvae. In addition, half of the plots were on a relatively fertile soil type, and the other half were on a relatively infertile soil type. Natural light levels in the plots were also varied such that half of the plots were assigned to a high-light treatment and half to a low-light treatment. Dyer and Letourneau maintained these treatments for 18 months and measured herbivory and leaf production within each of the plots.
If the production of the Piper trees was limited primarily by resource supply (bottom-up control), then the addition of the Tarsobaenus beetles would be expected to have little effect on Piper leaf production. Soil fertility and light levels would be expected to have greater effects on leaf production if these effects were more important than the influence of the trophic cascade associated with beetles, ants, and herbivores (top-down control). Dyer and Letourneau found, however, that the trophic cascade was the only significant influence on leaf production.
The addition of the predatory beetles decreased ant abundance fivefold, increased rates of herbivory threefold, and decreased leaf area per tree to half that in the control plots (FIGURE 21.12). This experiment provided convincing evidence of a trophic cascade affecting the production of the Piper trees. It should be noted, however, that the lack of an effect of soil fertility and light in the control treatments, which had low rates of herbivory, indicates that the resource(s) that actually limit production may not have been manipulated in this experiment. An additional experiment that used more controlled manipulation of light levels and soil nutrients, rather than relying on variation in natural levels, found significant effects of these resources on Piper production, but it also found a continued strong effect of herbivory (Dyer and Letourneau 1999b). Thus, it is clear that trophic cascades do occur in diverse terrestrial ecosystems, although they may require strong interactions between specialist predators and their prey.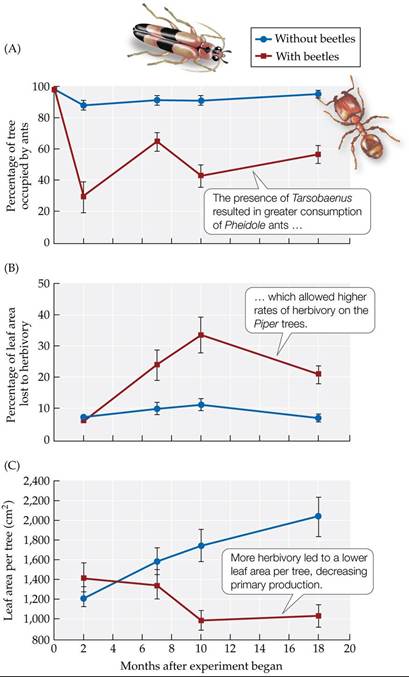
FIGURE 21.12 Effects of a Trophic Cascade on Production Atrophiccascadeina tropical rainforest understory ecosystem (see Figure 21.11) was shown to have important effects on (A) predation, (B) herbivory, and (C) production. Error bars show ± one SE of the mean. (After L. A. Dyer and D. K. Letourneau. 1999a. Proc NatlAcad Sci USA 96: 5072-5076. © 1999 National Academy of Sciences, U.S.A.) View larger image
More on the topic Trophic interactions can trickle down through multiple trophic levels:
- CONCEPT 21.3 Changes in the abundances of organisms at one trophic level can influence energy flow at multiple trophic levels.
- What determines the number of trophic levels?
- Organisms can be grouped into trophic levels
- Energy flow between trophic levels differs among ecosystem types
- Energy flow between trophic levels can be depicted using energy or biomass pyramids
- Energy Flow between Trophic Levels
- The strengths of trophic interactions are variable
- CONCEPT 21.1 Trophic levels describe the feeding positions of groups of organisms in ecosystems.
- CONCEPT 21.4 Food webs are conceptual models of the trophic interactions of organisms in an ecosystem.
- Trophic efficiencies can influence population dynamics
- Trophic Cascades
- The way we think about species interactions changes dramatically when we consider that they are embedded in a community of multiple interactors.