Retroelements and retrovirus INFECTIONS
Retroviruses are the phylogenetic pinnacle of a diverse assemblage of related entities known collectively as ret- roelements, which occupy over 37% of the mouse genome. Most of these elements can be considered genetic parasites or fossil DNA, but they play a significant role in laboratory mouse strain characteristics and disease.
Despite their significance, retroelements are often unfortunately ignored in texts on mouse biology. The reader is therefore herein subjected to an overview of retrovirus biology in order to better appreciate the role of retroelements in pathogenesis of disease and phenotypes in the laboratory mouse.The mouse genome is replete with literally thousands of copies of endogenous integrated retroelements that require reverse transcription from RNA to DNA. Among the retroelements are replication-competent endogenous retroviruses whose genomes encode gag, pro, pol, and env genes that are flanked on their 5'- and 3'- ends by long terminal repeats (LTRs). Endogenous retroviruses include murine leukemia viruses (MuLVs) and murine mammary tumor viruses (MMTVs), which are genetically unrelated. MMTVs have a slightly longer genome than MuLVs, and their LTR regions encode a critically important gene: Sag (superantigen). Endogenous MuLVs and MMTVs are autonomous, in that they encode their own reverse transcriptase. Endogenous retroviruses are integrated as DNA within the genome, wherein they are termed "proviruses." The majority of endogenous retroviruses (proviruses) are defective, ranging from being one base pair substitution away from an active provirus to having multiple stop codons, frameshifts, and deletions that render them incapable of expression as a replication-competent virus. Furthermore, most proviruses are methylated and transcriptionally silent. Other autonomous retroelement families that are flanked by LTRs include intracisternal A particles (IAPs), MusD elements, VL30 elements, glutathione tRNA primer binding sites (GLNs), and murine endogenous retroele- ments (MuERVs), which include MuERVC, murine retrovirus-related sequences (MuRRS), and murine repeated viruses on Y chromosome (MuRVYs).
These elements are missing significant parts of their genome (especially env) or encode no open reading frames at all. They are related to retroviruses because of similarities within their LTRs, and each family of retroelements has its own unique LTR sequences. There are also autonomous retroelements that lack LTRs, known as long interspersed nucleotide sequences (LINEs), which make up 20% of the mouse genome. Related retroelements that are nonautono- mous, and therefore must borrow reverse transcriptase function from autonomous retroelements, are termed "retrotranscripts." These elements encode no proteins, but are usually flanked by LTRs. They include early transposons (ETns) and short interspersed nuclear elements (SINEs), which make up 8% of the genome. Finally, there are numerous solo LTRs, which are devoid of internal reading frames.Replication-competent, endogenous MuLVs and MMTVs are closely related to exogenous MuLVs and MMTVs, which are not integrated within the mouse genome. Exogenous retroviruses are transmitted horizontally as conventional viruses. Exogenous viruses exist within wild mouse populations, but they have been eliminated (unless purposely reintroduced) from laboratory mice by cesarean rederivation and/or foster nursing. The well-studied Friend, Moloney, and Rauscher (FMR) group of MuLVs and the Bittner milk agent (MMTV-S) are exogenous retroviruses. The small number of replication-competent endogenous viruses can also be horizontally transmitted to other mice, if the mice possess the appropriate susceptibility factors.
Retroviruses and retroelements are highly promiscuous. Retroviruses possess a diploid genome, which incorporates two RNA genome strands. Virion assembly within host cells that are coinfected with either exogenous or endogenous retroviruses often results in acquisition of 2 different viral genomes within a virion. Reverse transcriptase readily jumps from homologous regions of one strand to the other, thereby creating recombinant forms of viral genomes.
In this way, otherwise defective endogenous retroviruses, and even retro- elements such as MuRRS and ETns, can contribute to generation of new replication-competent recombinant forms of virus with altered biological behavior. Some retroelements, such as VL30s, are frequently incorporated as passengers (parasitic RNA) during MuLV virion packaging and have a high recombination rate with retroviruses. Although most endogenous retroelements are defective, they represent mobile DNA species that can reintegrate in other regions of the genome during cell division without virion assembly and re-infection. These are known as retrotransposons.Occasionally, retroviral integrations within the host genome result in usurping components of the host DNA, which becomes incorporated in the viral genome. These regions of host DNA tend to result in loss of critical viral sequences, and the virus becomes defective. Acute transforming MuLVs and murine sarcoma viruses (MuSVs) are viruses that have incorporated a host cellular protooncogene (c-onc) that directly alters cell division. Once in the viral genome, the c-onc is referred to as viral oncogene (v-onc). Often, v-oncs are mutated, which increases their pathogenicity. Acute transforming retroviruses are so named because they carry their own v-onc that induces rapid transformation and evolution of neoplasia (in contrast to chronic retroviruses that induce neoplasia by random insertional mutagenesis). Examples of acute transforming retroviruses are Abelson MuLV (the v-onc is abl) and Moloney MuSV (the v-onc is mos). Since acute transforming MuLVs and MuSVs are experimental tools, and do not appear to contribute to natural disease, they will not be discussed further. Defective proviruses, including acute transforming MuLVs and MuSVs, can be rendered infectious through assistance of helper viruses, which contribute the missing structural elements for virion assembly, particularly envelope proteins, and/or LTR elements that confer tissue specificity.
Each inbred strain of mouse has its own characteristic "signature" of endogenous retrovirus and retroelement sites of integration in its genome. Related mouse strains share similarity in these patterns of integrated proviruses. Furthermore, different families of retroviruses have contributed to the mosaicism of the laboratory mouse genome from different Mus species and at different times during the evolution of the mouse. Based upon genome sequence comparison among Mus species, retrovirus-like elements, such as Etns, probably entered the mouse genome 5-10 million years ago, whereas MuLVs and MMTVs entered the genome within the last 1.5 million years and are still in the process of entry in some cases. A progressive chronology of integration helps to explain retrovirus biology. Older elements represent “fossil” DNA with sequences that are quite divergent from more recent families of retrovirus. They tend to be highly methylated and have major sequence deletions, stop codons, frameshifts, and other characteristics that make them defective. The most recent acquisitions are the MuLVs and MMTVs, which also may exist in multiple copies throughout the genome, most of which are also transcriptionally silent or defective. Only a relative few are transcribed as replicating viruses capable of infecting other cells. These are the viruses that will be emphasized, but it must be recognized that even defective retrovirus-like elements can become reintegrated within the dividing cell genome as retrotransposons. This process takes place within the nuclear context of the dividing cell, in contrast to replicating viruses, which leave the cell and enter another. This explains the acquisition of multiple copies of these older elements throughout the genome. For example, there are 100-200 copies of VL30s, and 1,000-2,000 copies of ETns in the mouse genome that have arisen from retro- transposon activity.
Retroviruses of the mouse can best be described as “mother nature's transgenes,” with a major component of their biology involving integration within the genome of dividing cells through viral reverse transcriptase.
When integration involves somatic cells, the outcome can be variable, but integration in germ cells results in passage of the provirus within the genome to subsequent mouse generations. Integration is random and usually phenotypically silent, but integration in other sites can result in insertional mutagenesis. Retro- elements and their mouse host have evolved a remarkable state of detente. Endogenous retroviruses have undergone adaptations that render them relatively innocuous to their hosts. They tend to be non- pathogenic and replicate poorly. Many show their phylogenetic age and are defective and transcriptionally silent. The host has also evolved to cope with their presence by mutation or loss of key viral receptors and other factors, thereby precluding reinfection (and reintegration in other sites of the genome that may result in deleterious effects).Finally, an introduction to mouse retrovirus biology is not complete without understanding virus tissue and host species tropism. LTRs contain coding sequences, including enhancers and promoters, which dictate transcriptional activity and tissue specificity. The MMTV LTR, for example, confers tropism for mammary tissue, which is why MMTV LTR is often used in transgenic constructs for mammary cancer research. Endogenous MuLVs are generally classified biologically by their host species tropism. Ecotropic MuLVs have tropism for mouse cells, but not cells of other species. Furthermore, ecotropic viruses display differential tropism for cells of different mouse strains, through allelic variation of the Fv-1 gene. Fv-1 has 2 major allelic variants, Fv-1n and
Fv-1b, that are codominant. Mice that are homozygous for Fv-1n are permissive for infection with N-tropic ecotropic viruses, whereas mice that are homozygous for Fv- 1b are permissive for infection with B-tropic ecotropic viruses, and Fv-1b/n heterozygous mice are permissive to both types of ecotropic virus.
Fv-1n mouse genotypes include AKR, CBA, C3H, C57L, C57BR, C58, and SWR strains, among others. Fv-1b mouse genotypes include A, BALB/c, C57BL/6, C57BL/10, FVB, and others. A third Fv-1 allele, Fv-1nr, is found in AKR, NZB, NZW, and RF mice. Mice that carry ecotropic MuLV can effectively control ecotropic infection through the Fv-1 locus. For example, BALB mice carry an endogenous N-ecotropic virus, but it cannot replicate (and re-infect and re-integrate) within BALB mice. Xenotropic MuLVs have tropism for cells of other species but not mouse cells. This is an apparent example of the mouse evolving away from susceptibility to these endogenous viruses, as mice have lost or mutated the functional cellular receptor. Polytropic viruses have tropism for mouse, as well as other species. Based upon LTR sequences, the polytropic group has two subgroups: polytropic and modified polytropic viruses. Both have LTR insertions derived from MuRRS, but all endogenous polytropic viruses are defective.Additional types of MuLV and retroelements exist in wild, but not laboratory, mice. A fourth group of viruses, known as amphotropic viruses, have tropism for cells of mice and other species, but they differ from polytropic viruses in their LTR sequences and receptor specificity and represent a family of viruses that are found in wild, but not laboratory, mice. It is generally believed that the Friend, Moloney, and Rausher MuLVs are related to the amphotropic group. Wild mice may also have exogenous ecotropic MuLV that is unrelated to the endogenous ecotropic viruses in laboratory mice. Fv-4 is another extensively studied resistance determinant for MuLV replication, but it has been found only in wild mice and is apparently not represented in the strains of laboratory mice that have been examined. It, like Fv-1, is derived from retrovirus sequences.
Ecotropic proviruses represent the most recent acquisitions into the mouse genome, and their presence varies among laboratory mouse strains. Several mouse strains, including 129, NZB, NFS, C57L, SWR, and CBA, are ecotropic virus-negative. Others, including BALB/c, A, C3H/He, and CBA, possess a single ecotropic virus, with low expression, and C57BL/6, C57BL/10, and C57BR possess a different, single ecotropic virus. A few exceptional mouse strains, most notably AKR, C58, and HRS, have multiple genome-length ecotropic proviruses, with high levels of expression. Ecotropic proviruses are given gene names (Emv-1, Emv-2, etc.). The multicopy Emv, formerly called Akv, that is present in AKR, C58, and HRS but not other strains of mice is a genetic contribution from M. molossinus. Xenotropic viruses are present in multiple copies in all mouse strains, but with generally low expression levels as replication-competent virus, with one exception being the NZB mouse, which expresses high levels of xenotro- pic virus in its tissues. Xenotropic proviruses are given the gene designation Xmv (Xmv-1, Xmv-2, etc.). Polytropic retroviruses, which probably represent the oldest of the endogenous retrovirus family in terms of germline acquisition, are present in all strains of mice, but all polytropic viruses are defective. Polytropic provirus genes are named Pmv and modified polytropic provirus genes are named Mpmv. There are over 50 MMTV provi- ral loci, designated Mtv-1 to Mtv-56, in various strains of mice. The majority of these proviruses do not produce infectious virus, with the notable exception of Mtv-1, which is present in DBA and C3H mice, and Mtv-2, which is present in GRS mice. Every chromosome of the mouse genome has multiple endogenous Emv, Xmv, Pmv, Mpmv, or Mtv proviruses. Some populations of wild mice have been found that lack both exogenous and endogenous MMTV.
Germline Insertional Mutagenesis
Reintegration of endogenous retroviruses or transposition of retroelements can and do result in spontaneous mutations among inbred mice. Examples of retrovirus integrations that have resulted in spontaneous mutations include the dilute (d) mutation of DBA mice due to insertion of an ecotropic virus into the Myo5a locus, the hairless (hr) mouse mutation due to a polytropic virus integration in the hr locus, and the rodless retina (rd1) mutation due to integration of a xenotropic virus into the Pde6b locus. Germline integrations of IAPs and ETns have also contributed to nearly 15% of all known spontaneous mutations, resulting in a wide variety of overt phenotypes, including athymia (Foxn1), stargazer (Cacng2), obese (Lep), and albino (Tyr), among many others. Like transgenes, spontaneous phenotypic rever- tants can occur when the offending retrovirus element is excised by DNA repair. Retrovirus integrations are usually random (although there are “hot spots” within the genome), and the resulting integrated DNA within the genome is “hemizygous,” analogous to a transgene. Ecotropic viruses are responsible for the greatest number of novel proviral integrations in the germline. The artificial process of inbreeding laboratory mice has made these hemizygous integrations homozygous and thus they have been given gene names. Examination of the genomes of various substrains derived from parental strains has revealed both acquisition and loss of proviruses over the course of recent mouse history, contributing significantly to substrain genetic divergence and emphasizing the dynamic nature of retroviruses and retroelements.
Modulation of the Host Immune Response by MuLVs and MMTVs
Endogenous retroviruses and retroelements that encode proteins during the life of a mouse may elicit varying degrees of host immunity. Viruses that are expressed in high titer and in all tissues during early development are likely to induce immunological tolerance, particularly if expressed in the thymus. The superantigens (SAgs) of some MMTVs stimulate and deplete specific Vβ T-cell subsets (see “MMTVs,” later in this chapter). However, endogenous retroviruses or retroelements may not be expressed in tissues until later in life, and thus are likely to stimulate a host immune response, albeit weak. Proviruses may be transcriptionally silent until there is a stimulus for cell division, at which time they may be expressed and stimulate a host response. Thus, the host's immune response to its load of endogenous retroviruses is variable, but often results in the evolution of immune complexes as mice age that are suspected to contribute to the development of spontaneous vasculitis and glomerulonephritis. These disease syndromes, not surprisingly, vary in severity among different mouse strains. Expression of endogenous retroviruses in pancreatic islets of NOD mice is associated with the evolution of immune- mediated insulitis, and some have speculated that retroviruses may contribute to autoimmune disease in NZB x NZW hybrid mice.
Murine Leukemia Viruses
Exogenous MuLVs and replication-competent endogenous MuLVs are transmitted through milk and to a lesser extent semen, saliva, venereal transmission, perinatal infection, or transplacental transmission. Germline integration by MuLVs tends to be primarily through infection of dividing ova rather than male gametes. Reintegration of MuLVs into the genome of somatic cells results in random insertional mutagenesis, and neoplasia is favored when integration takes place near host proto-oncogenes.
The most extensively studied system for endogenous MuLV pathogenesis has been the AKR mouse (also C58 and HRS strains). AKR mice develop nearly 100% thymic lymphomas of T-cell origin within 6-12 months of age (Fig. 1.44). The AKR mouse genome contains over 40 provirus integrations, including 3 critical parental proviruses. All tissues express high levels of replication- competent ecotropic virus (Emv-11) early in life. This virus is B-tropic and incapable of infecting AKR cells, which are Fv-1n/n homozygous. However, high-level expression in cells that are coexpressing xenotropic virus results in genetic recombinant viruses with altered LTR sequences derived from the xenotropic parent. These recombinant viruses are expressed in cells that also coexpress a polytropic virus, which donates altered envelope (env) sequences and allows infection of thymic cells. These recombinant viruses undergo duplication of enhanced LTR sequences, with high levels of replication in the thymus, which favors insertional mutagenesis. The proximal viral pathogen, which represents a recombinant of 3 parental endogenous ecotropic, xenotropic, and polytropic parents, possesses the ability to induce foci in mink cells. It is thus called a mink cell focus (MCF)
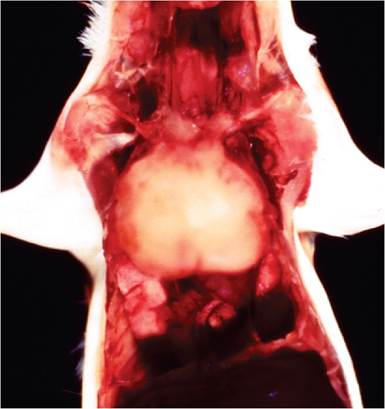
FIG. 1.44. Thymic lymphoma in an AKR mouse due to naturally occurring recombination of endogenous retroviruses. Nearly all mice of this strain develop thymic lymphoma between 6 and 12 months of age.
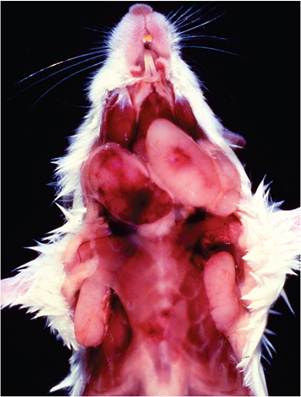
FIG. 1.45. Multicentric lymphoma in a BALB/c mouse due to naturally occurring recombination of endogenous retroviruses. Note the markedly enlarged cervical and axillary lymph nodes.
forming virus. MCF viruses are the product of in vivo recombination and do not exist as natural germline proviruses. These events are highly artificial laboratory mouse consequence from selective breeding for the thymic lymphoma phenotype and inbreeding. There is no evidence for MCFs among wild mice.
The majority of mouse lymphomas are of B or pre- B-cell origin and tend to arise in the spleen. They arise in 40% of NFS mice, which lack ecotropic virus, but with a longer latency period and lower grade than mice carrying ecotropic virus. Neonatal thymectomy of AKR mice results in B-cell lymphomas arising from the spleen, and congenic mice derived from ecotropic virus-free NFS mouse background that have been backcrossed with the AKR Emv provirus locus develop splenic B-cell tumors. B-cell tumors with ecotropic virus integrations result from random integration of the ecotropic virus and are not necessarily associated with activation of host oncogenes. The virus is believed to be functioning to increase the rate of clones and speed the evolution of lymphomas that would otherwise arise but at a slower pace. BALB/c mice develop a late-onset lymphoma that is multicentric (Fig. 1.45). They carry nononcogenic, N-ecotropic, and xenotropic viruses that form recombinants that are oncogenic and B-ecotropic, resulting in infection of lymphoid cells, integration, and late-onset lymphoma. Each mouse strain possesses its own set of proviruses and host factors, with varying degrees of MuLV expression and disease that are mouse strain-, age-, tissue-, and cell type-specific.
Not all outcomes of MuLV infection are neoplastic, and in fact most are phenotypically silent. Other reported syndromes include altered coat color and consistency, hairlessness, central nervous system disease (see Arterivirus, LDV Infection in this chapter), premature graying, and potentially any other phenotype resulting from random integration or infection with expression of viral gene products. The most widely studied MuLV-related neurologic syndrome has been observed in feral Lake Casitas mice, which naturally develop hind limb paralysis. The syndrome is caused by MuLV CasBr-E, an ecotropic virus that is distinct from ecotropic MuLVs of laboratory mice. Disease is readily reproduced by inoculation of Fv-1n/n laboratory mice. Diseased mice manifest neuronal loss with proliferation and hypertrophy of microglia, vacuolation, and spongiosis, particularly in the ventral horns of the lumbar spinal cord.
Murine Mammary Tumor Viruses
Although MMTVs are generally associated with mammary gland neoplasia, their lymphocytotropism and ability to transform lymphoid cells through insertional mutagenesis is an equally important aspect of their biology. Like MuLVs, MMTVs are either exogenous or endogenous. Exogenous and replication-competent endogenous MMTVs are transmitted primarily in the milk, and to a lesser extent saliva. Exogenous MMTV, which is known as MMTV-S (Standard), the “extrachro- mosomal milk factor,” or the “Bittner agent,” has been eliminated from modern mouse populations by cesarean rederivation or foster nursing, unless intentionally maintained for experimental purposes. In contrast, all strains of laboratory mice possess varying numbers of endogenous MMTV provirus (Mtv) in their genome. All exogenous and some endogenous MMTVs are expressed in mammary tissue and lymphoid tissue, but some endogenous MMTVs, including Mtv-7 and Mtv-9, are not expressed in mammary tissue. Like MuLVs, most MMTV endogenous proviruses are defective, but can recombine with replication-competent MMTVs.
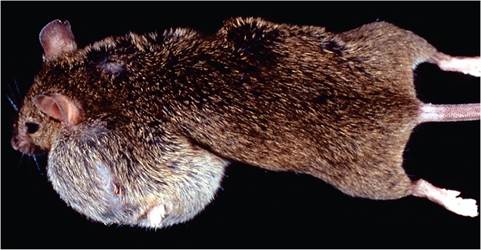
FIG. 1.46. Mammary neoplasia in a C3H mouse due to naturally occurring expression ofendogenous mouse mammary tumor virus (MMTV).
As their name implies, MMTVs are associated with mammary neoplasia through random integration. Genetically susceptible strains of mice, such as C3H, infected with exogenous MMTV-S, develop a high prevalence and early onset of mammary tumors. Mice that carry replication-competent endogenous Mtv-1 (C3H, DBA) or Mtv-2 (GRS) also develop mammary neoplasia (Fig. 1.46), but onset is delayed. MMTV dual lymphoid and mammary tropism, enhanced by LTR factors, favors this event. Exogenous MMTV (and replication-competent endogenous MMTV) is transmitted primarily from infected dams to nursing pups through the milk. Virus initially infects gut-associated M cells and dendritic cells and then replicates in B cells of Peyer's patches. Infected B cells express viral LTR-encoded superantigen (SAg) in the context of MHC class II on T cells, which results in stimulation and proliferation of T cells through recognition by specific Vβ T-cell receptors. Activated T cells release lymphokines that further stimulate proliferation of bystander B cells. Dividing B cells are requisite for retrovirus infection, thereby amplifying virus- infected B lymphocytes, which subsequently transport MMTV to mammary tissue. The role of SAg is critical in MMTV biology, as the more the virus produced, the greater the efficiency of transmission in the milk. Once virus enters the mammary gland via infected lymphocytes, it further replicates extensively within mammary tissue. Mammary tropism is favored by promoters and enhancers in the LTR region, thereby amplifying virus and favoring transmission in the milk. High titers of the virus within mammary tissue increase the chance of proviral integration near a site of a cellular protooncogene, with subsequent oncogenesis. There is a high correlation between virus titer in milk and mammary tumor incidence within a mouse strain. MMTV also utilizes other mechanisms that favor its replication by circumventing the host innate immune response.
In contrast to mice exposed to exogenous MMTV, mice that express SAg from an endogenous MMTV develop selective deletion of the corresponding Vβ T-cell subset. These mice resist infection with exogenous MMTV with the same SAg specificity, since the exogenous virus cannot effectively amplify itself in the Peyer's patches. In addition, Vβ6 T-cell depletion in mice with endogenous Mtv-7 is a major factor for polyoma virus-induced neoplasia susceptibility in C3H/ Bi, C58, CBA, AKR, and RF mice. Mice that are resistant to polyoma virus neoplasia, such as C57BR, C3H∕He, and CBA, lack Mtv-7, but possess other Mtvs with different SAg specificity. Lack of appropriate Vβ6 T-cell subset, induced by Mtv-7 SAg, precludes host T-cell immunity against polyoma virus-induced tumors.
Alternatively, MMTV proviruses may not be expressed until later in life, so there is no early Vβ T-cell deletion. SJL mice, for example, carry endogenous Mtv-29 that is initially transcriptionally silent but expressed later in life when Peyer's patches are stimulated antigenically. Over 90% of SJL mice develop MMTV-associated lymphoma by 13 months of age. Expression of SAg results in massive T-cell-mediated B-cell proliferation, with B-cell lymphoma arising in Peyer's patches, mesenteric lymph nodes (Fig. 1.47), and subsequently other sites. The ensuing follicular center cell lymphoma is characteristic of this strain of mouse. A similar pattern of follicular center cell lymphoma arising from gut-associated lymphoid tissue is seen in other strains of mice, but later in life. For example, C57L mice also express Mtv-29, but suppress development of lymphomas by NK cells, whereas SJL mice develop a high prevalence and earlier onset of lymphomas because they are NK cell-deficient. MMTV can also be associated with T-cell lymphomas of the thymus in GR and other strains of mice. MMTVs derived from these
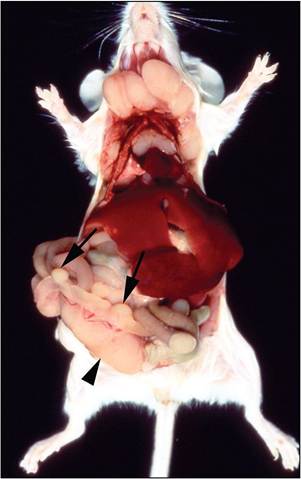
FIG. 1.47. Multicentric lymphoma arising from gut-associated lymphoid tissue in a SJL mouse due to naturally occurring expression of endogenous MMTV. Note the prominent Peyer's patches (arrows) and enlarged mesenteric lymph node (arrowhead).
tumors have undergone a change in cell tropism from B cells to T cells, which is mediated through rearrangements in their LTR regions.
Diagnosis
Diagnosis of MuLV or MMTV is not necessary, since all mice are infected or have provirus sequences within their genome. Electron microscopic examination of normal and neoplastic mouse tissues frequently reveals C- type (MuLV), A-type (IAP), and B-type (MMTV) particles as an incidental findings.
Significance
MuLVs and MMTVs are an integral and important part of mouse biology and inseparable features of the laboratory mouse. They are valuable models for retrovirus pathogenesis. They become significant as natural pathogens when they induce life-limiting disease and cause spontaneous mutations and substrain divergence. For example, the emergence of an ecotropic MuLV in a colony of CFW Swiss mice has resulted in a high incidence of lymphoma in a stock that was previously characterized as having a low incidence of spontaneous lymphoma. The consequences of recombinant retroviruses arising from microinjection of ES cells into blastocysts of different mouse strain background have not been fully realized in chimeras and their progeny. MuLVs are known to recombine with HIV in SCID-hu mice, which limits the value of this model.
More on the topic Retroelements and retrovirus INFECTIONS:
- AVIAN RETROVIRUS INFECTIONS
- OTHER RETROVIRUS INFECTIONS
- CHAPTER 15 RETROVIRUS INFECTIONS
- Other Bacterial Infections Corynebacterium spp. Infections
- Bloodstream Infections and Catheter-Related Bloodstream Infections
- Skin, Soft-Tissue, and Bone Infections Purulent Skin and Soft-Tissue Infections (Furuncles, Carbuncles, Abscesses)
- OPPORTUNISTIC INFECTIONS
- INTRAUTERINE INFECTIONS
- PERINATAL INFECTIONS
- NOSOCOMIAL INFECTIONS
- INTRACRANIAL INFECTIONS
- Other Virus Infections of Minimal
- ENTERQViRAL INFECTIONS (NON-POLIO)
- Genitourinary Infections
- HERPETIC VIRAL INFECTIONS
- FUNGAL INFECTIONS
- Viral infections
- Tick-Borne Infections