Imports and upwelling are important sources of nutrients in marine ecosystems
Rivers join marine ecosystems in estuaries (described in Concept 3.3). In these zones where fresh water meets seawater, salinity—and thus water density—is variable. This variation influences the mixing of waters and the chemical forms of some nutrients.
For example, phosphorus bound to soil particles may be released in a form more easily available to phytoplankton as a result of changes in pH when river water mixes with seawater.As the velocity of water flow decreases toward the mouth of a river, suspended sediments begin to settle out of the water. These sediments are substrates for detritivores and nutrients for phytoplankton in the estuary. Estuaries are often associated with salt marshes, which are rich in nutrients because they trap both riverine and ocean sediments. Like benthic sediments in lakes, estuarine and salt marsh sediments have low oxygen concentrations that limit decomposition.
As described in Concept 20.2, primary production in the open ocean is limited by several nutrients, including nitrogen, phosphorus, and, in some areas, iron and silica. Seawater has relatively high concentrations of magnesium, calcium, potassium, chloride, and sulfur. Sources of nitrogen in marine ecosystems include inputs from rivers and atmospheric deposition as well as tight internal cycling through decomposition. Rates of nitrogen fixation by cyanobacteria in the oceans are lower than those in freshwater lakes, possibly because these organisms are limited by molybdenum, which is a component of the nitrogenase enzyme. Phosphorus, iron, and silica enter the marine ecosystem primarily in dissolved and particulate form in rivers; a smaller but important contribution comes from atmospheric deposition of dust. Inputs from both of these terrestrial sources are increasing as a result of human activities, including large-scale desertification and deforestation.
Deep deposits of sediments (up to 10 km, or 6 miles thick!) have accumulated in the benthic zones of the open ocean. These deposits, which consist of a mix of ocean-derived detritus and terrestrial erosional sediments, are important potential sources of nutrients. Sulfate reduction and denitrification occur in these anoxic sediments, and some decomposition and mineralization of organic matter also occur there. Bacteria have been found as deep as 500 m in these sediments. The deep ocean layers are dense due to cold temperatures and high salt concentrations (see Concept 2.2) and usually don't mix with the surface waters. Mixing of deep, nutrient-rich waters with nutrient-poor surface waters does occur in zones of upwelling, where ocean currents bring deep waters to the surface (FIGURE 22.19). These zones of upwelling are highly productive and thus are important areas for commercial fisheries.

FIGURE 22.19 Zones of Upwelling Enhance Nutrient Supply for Marine Ecosystems Phytoplankton blooms (green areas), fed by upwelling of nutrient-rich deep ocean water, can be seen off the coast of the Pribilof Islands (Alaska) in this satellite image. View larger image
A Case Study Revisited
A Fragile Crust
We've seen that nutrient supplies for plants in terrestrial ecosystems are dependent on the weathering of rock minerals and the decomposition of detritus in the soil, as well as on the fixation of atmospheric nitrogen. How might the loss of biocrusts from desert soils influence these processes? As this chapter's Case Study explained, the crusts prevent erosional losses of soil by helping to bind soil particles together. The activity of the organisms that make up the crusts may also influence nutrient inputs and, in turn, the productivity of the desert ecosystem, as well as its capacity to withstand the desert climate.
Jason Neff and colleagues conducted a study to evaluate the effects of cattle grazing on soil erosion and nutrient availability on the Colorado Plateau (Neff et al.
2005). They selected three study sites in Canyonlands National Park: one that had never been grazed and two that had been grazed historically but were closed to grazing after 1974 (30 years of recovery). Cattle grazing in the park first occurred in the 1880s, and most of its soil surface has been affected. The ungrazed study site was surrounded by rock formations that prevented the movement of cattle into the area. The study sites all had the same parent material and similar plant communities and were located within 10 km of one another. Biocrusts were present at all three sites, although those at the historically grazed sites had clearly been damaged, as they appeared less well developed than those at the site that had never been grazed.Samples of soil and bedrock were collected from each of the sites, and the textures and nutrient contents of the soils were compared. In addition, the retention of fine dust from the atmosphere was estimated by measuring the magnetic properties of the soil. Dust blown in from distant areas contains higher amounts of iron oxides than the native soil, so the more dust present, the stronger the magnetic signal. Retention of this dust is important because it is a source of mineral nutrients. In addition, loss of this dust indicates the potential for erosional loss of the native soil as well.
Neff and colleagues found that the historically grazed soils had less fine-textured soil, and substantially less magnesium and phosphorus, than the ungrazed soils (FIGURE 22.20). They attributed these differences to greater retention of dust and lower loss of native soils with better-developed biocrusts. The crusts may also enhance rates of weathering by altering pH, by increasing the rates of chemical reactions that release mineral nutrients, and by increasing water retention in the soil. Soils in the historically grazed sites also contained 60%-70% less carbon (from organic matter) and nitrogen than those in the ungrazed sites. These differences were also related to the development of biocrusts.
Although a crust had begun to recover at the historically grazed sites, comparison with the ungrazed site showed that the cumulative loss of carbon and nitrogen from the soils during the period of grazing was high. The cyanobacteria in biocrusts fix atmospheric N2 (Belnap 2003), which represents an important input of a nutrient that may limit plant growth in the absence of water limitation during the spring growing season. In addition, crust-covered soils absorb more solar radiation and retain more water than soils without crusts, creating conditions more conducive to decomposition and mineralization.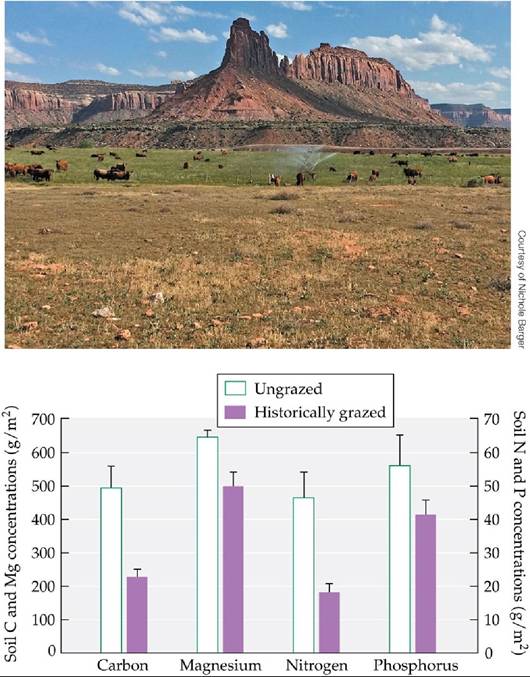
FIGURE 22.20 Loss of Biocrusts Results in Smaller Nutrient Supplies Historically grazed soils in Canyonlands National Park contained less carbon, magnesium, nitrogen, and phosphorus than soils that had never been grazed. Error bars show one SE of the mean. (Graph after J. C. Neff et al. 2005. EcolAppl 15: 87-95.) View larger image
Connections in Nature
Nutrients, Disturbance, and Invasive Species
By increasing nutrient supplies and stabilizing soils, biocrusts enhance primary production. Plants growing in association with the crusts have higher growth rates, and contain more nutrients, than plants growing in soils without crusts. Plant cover also increases in the presence of biocrusts. Furthermore, biocrusts have been shown to lower the germination and survival rates of invasive plants (Havrilla et al. 2019) (see Chapter 23). Thus, the destruction of crusts by cattle grazing has had multiple ecological effects.
Are the negative effects of cattle grazing on soil stability and nutrient availability that Neff and colleagues observed in Canyonlands National Park common in other areas? The answer lies in part with the long-term history of grazing and climate in North America. Prior to Euro-American settlement, soils in much of the intermountain West did not experience the amount of grazing by native animals that occurred in other areas, such as the Great Plains, where large herds of bison roamed (see the Case Study in Chapter 3 and the discussion in Concept 3.2).
A combination of aridity and long-term development of biocrusts may have given the soils of the Colorado Plateau an especially low tolerance for heavy grazing.In the grasslands of the intermountain West, the combination of soil disturbance and loss of biocrust has created a situation conducive to the spread of non-native species—most notably cheatgrass (Bromus tectorum; FIGURE 22.21), a native of Eurasia. Cheatgrass has had profound effects on the ecology of much of western North America. Cheatgrass is a spring annual that sets seed, dies, and dries out by early summer. This life history increases the amount of dry, combustible vegetation that is present during the summer. As a result, cheatgrass has increased the frequency of fires, which now occur about every 3-5 years, compared with more natural fire frequencies of 60-100 years. Native grasses and shrubs cannot recover from such frequent fires, so cheatgrass increases its dominance under these conditions. Cheatgrass is an effective competitor for soil resources, and it also lowers rates of nitrogen
cycling by producing litter with a C:N ratio higher than those of native species (Evans et al. 2001). This combination of increasing fire frequency, increasing competition, and changes in nutrient cycling has led to decreases in native species richness in many parts of the intermountain grasslands.

FIGURE 22.21 ScourgeoftheIntermountainWest Large areas of the intermountain West of North America are now dominated by cheatgrass (Bromus tectorum), an invasive species that increases fire frequencies, outcompetes native plants for resources, and spreads rapidly across the landscape. View larger image
More on the topic Imports and upwelling are important sources of nutrients in marine ecosystems:
- Catchment studies measure losses of nutrients from ecosystems
- Nutrients in Aquatic Ecosystems
- Minerals and atmospheric gases are the ultimate sources of nutrients
- CONCEPT 22.3 Nutrients cycle repeatedly through the components of ecosystems.
- Sources of Marine Pollution
- CONCEPT 22.4 Freshwater and marine nutrient cycles occur in a moving medium and are linked to terrestrial ecosystems.
- CONCEPT 22.1 Nutrients enter ecosystems through the chemical breakdown of minerals in rocks or through fixation of atmospheric gases.
- CONCEPT 22.2 Chemical and biological transformations in ecosystems alter the chemical form and supply of nutrients.
- Sources of!important; text-transform:uppercase'>Law from the Republic to the Dominate
- Nutrients in streams and rivers cycle while moving downstream
- Microorganisms modify the chemical form of nutrients
- Nutrients in lakes cycle efficiently in the water column
- Marine biological zones have been impacted by human activities
- Chapter 1 dealt with the main sources of Roman private law, in the sense of the formal sources which created it.
- CALICIVIRUS INFECTIONS OF MARINE MAMMALS
- Ecosystems
- Marine Pollution
- Nutrients cycle at different rates according to element identity and ecosystem type
- Marine Biological Zones