Helicobacter spp. Infections
Helicobacter spp. have emerged as a major group of gastrointestinal commensals with pathogenic potential in the laboratory mouse. Mice serve as hosts to several named species and additional novel isolates that are yet to be formally named.
Named species include H. hepa- ticus, H. bilis, H. muridarum, H. rodentium, H. typhlonius, H. ganmani, H. rappini, H. mastomyrinus, H. magdeburgen- sis, and H. pullorum. In addition, H. muricola has been isolated from the cecum of Korean wild mice (M.m. molossinus), and other novel species are continually being added to this list as they are isolated and characterized. Helicobacter spp. are microaerobic, curved to spiral rods with variable numbers of flagella. Each species has a somewhat distinctive electron microscopic appearance, which underscores their genetic (and antigenic) diversity. In the mouse, Helicobacter spp. occupy an enterohepatic niche and colonize the cecum and colon, with variable presence in the liver. Most immunocompetent mice develop minimal or no liver or intestinal lesions when infected with Helicobacter spp., but the pathogenic potential of these bacteria is manifest in a variety of immunodeficient types of mice. When disease occurs, typical lesions consist of hyperplastic typhlocolitis and hepatitis.There is evidence that H. bilis infections in humans may be linked to gall bladder, biliary and liver disease, and associated cancers, and H. pullorum is a human pathogen. Aside from their direct pathogenic potential, particularly in immunodeficient mice, the association of Helicobacter spp. with promotion of hepatic neoplasia and the presence of invasive and dysplastic intestinal lesions in infected mice have led to claims that Helicobacter is a carcinogen. Helicobacter hepaticus (and probably other Helicobacter spp.) is associated with an increase in hepatocellular tumors in certain strains of infected mice, and has also been shown to promote experimental chemical hepatocarcinogenesis.
Helicobacter typhlocolitis is extensively used as a model for human “inflammatory bowel disease” (IBD). Helicobacter typhlocolitis occurs in a variety of GEMs due to their various immunodeficiencies and is a classic example of “the disappearing phenotype syndrome” when GEM models are microbially rederived. The dysplastic and invasive nature of hyperplastic crypt epithelium, particularly when the mucosa is inflamed or eroded, may mimic neoplasia. These features, as well as proliferative kinetics (mitotic index and BUDR labeling patterns), lend credence to misinterpretation of lesions, with the use of terms like “carcinoma in situ” and “nonmetastatic colon cancer.” Similar lesions have been shown to arise in the colon of mice infected with C. rodentium that are totally reversible upon recovery. Although claims of neoplasia may or may not be valid, it is incumbent upon a pathologist who is familiar with mouse biology and biologic characteristics of neoplasia to accurately assess the validity of these lesions as bonafide neoplasia.Epizootiology and Pathogenesis
Natural outbreaks of hyperplastic typhlocolitis in immunodeficient mice have been associated with multiple species of Helicobacter, or (often) mixed infections. Observations by the authors have confirmed that most Helicobacter spp. of the mouse are capable of inducing proliferative typhlocolitis in immunodeficient mice, but some isolates may be nonpathogenic commensals. These agents appear to be widespread in conventional colonies of laboratory mice, but as awareness has arisen,
there has been a concerted effort to eliminate Helicobacter spp. from both commercial and research laboratory settings. Helicobacter hepaticus and H. rodentium appear to be the most prevalent members of the group among laboratory mice.
Mice are likely exposed to infection through ingestion of contaminated feces, and Helicobacter spp. are readily transmitted by contaminated bedding. Colonization of the cecum with H.
hepaticus has been associated with a significant decrease in the overall diversity of the cecal microflora. In B6 and A/JCr mice inoculated with H. hepaticus, the ceca were readily colonized with the organism. Although the ceca of B6 mice were more heavily colonized with H. hepaticus, significant lesions were, however, absent, and the inflammatory and immune responses were minimal compared with infected A/JCr mice. Helicobacter infections are persistent, with long-term fecal shedding of organisms. Transfer of neonatal mice from infected dams to Helicobacter- free foster dams has been found to be successful in eliminating the infection, providing the transfer occurred within the first 24 hours after birth. Cesarean rederivation and embryo transfer are effective means of eliminating infection. It is not safe to assume that infection is limited to intestine and liver in immunodeficient mice, as H. hepaticus has been documented to frequently contaminate transplantable human tumor xenografts when passaged through infected SCID mice. There is one report of transplacental infection of H. hepaticus in SCID/NCr mice.In the liver, H. hepaticus and H. bilis localize and persist indefinitely within the bile canaliculi. Mouse genotype is pivotal in expression of hepatic disease when infected with H. hepaticus. A/JCr, SCID/NCr, BALB/cANCr, C3H/HeNCr, and SJL/NCr mice have been found to be susceptible to hepatitis, whereas B6 and B6C3F1 mice are hepatitis-resistant. A/J mice also develop an earlier onset and higher prevalence of hepatocellular tumors when infected with H. hepaticus. Studies indicate that multiple genes are involved in genetic susceptibility and resistance to liver disease. Hepatic lesions are more common in male A/JCr mice than females, and the incidence of lesions is increased in mice 6 months of age or older. Other Helicobacter spp. may also be associated with hepatitis. For example, H. bilis has been associated with hepatitis in outbred Swiss Webster mice.
The pathogenesis of hepatitis and hepatocellular tumors is unknown, but hepatotoxins or autoimmunity to heat shock protein 70 are suspected. Colonization of the liver leads to induction of apoptosis and cellular proliferation, which are linked to promotion of neoplasia. Pathogenesis of typhlocolitis, which has marked inflammatory and hyperplastic features, has been studied in SCID and nude mice as well as GEMs with selective immune deficiencies, including IL-2, IL-10, T-cell receptor (alpha, beta, delta), RAG, MHC class II, and other null mutations. It has been proposed that Helicobacter infection induces a highly
Th1-polarized mucosal immune response, with production of IL-12, gamma interferon, and tumor necrosis factor (TNF) alpha in the lamina propria, resulting in expression of the epithelial mitogen, keratinocyte growth factor. Adoptive transfer of CD4 T cells expressing high levels of CD45RB into immunodeficient mice infected with H. hepaticus accelerates inflammation and hyperplasia, resulting in the conclusion that bowel disease is an immune-mediated disease that is directed against “normal” intestinal microflora.
Pathology
Infections in immunocompetent mice are usually inapparent. Clinical signs in immunodeficient mice include wasting (often complemented by pneumocystosis or other forms of pneumonia in immunodeficient mice) and mortality. Feces may be unformed, sticky, mucoid, or hemorrhagic. Rectal prolapse is a frequent, but nonspecific, sign of colitis in mice. The prolapsed mucosa is usually eroded and markedly inflamed and hyperplastic. These changes are nonspecific and due to the trauma of prolapse and may not be directly associated with Helicobacter spp. Thus, other areas of bowel should be examined to confirm the diagnosis. Segmental areas of cecum and colon are grossly thickened and opaque. Affected mucosa is thickened due to varying degrees of crypt hyperplasia, with markedly immature and mitotically active enterocytes occupying the entire crypt column.
Typical Helicobacter organisms can be readily demonstrated within crypt lumina of the affected sections of gut, using Steiner or other silver staining methods (Fig. 1.58). In severely affected mucosa,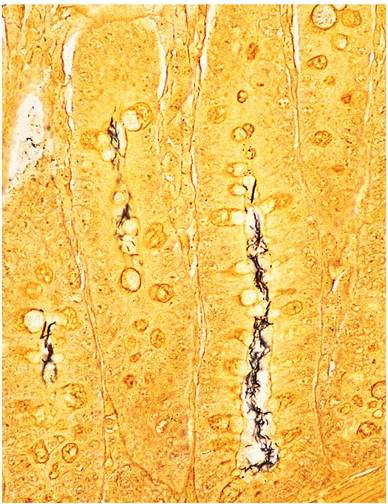
FIG. 1.58. Colon from an immunodeficient mouse, depicting Helicobacter organisms within hyperplastic crypts (Steiner stain).
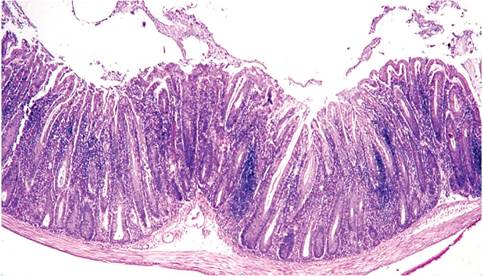
FIG. 1.59. Colon from a MRL-lpr (immunodeficient) mouse chronically infected with Helicobacter hepaticus. There is marked mucosal hyperplasia, with mononuclear cell infiltration in the lamina propria.
especially mucosa that has been aggravated by erosion or rectal prolapse, there is often focal crypt dysplasia and invasion of hyperplastic crypts into the underlying submucosa, accompanied by cystic change with mucous retention. Depending upon the mouse genotype and stage of infection, there are varying degrees of mixed leukocytic infiltration in the lamina propria. In some strains, there may be marked lymphocytic infiltration (Fig. 1.59). Although there may be extensive lesions in the large intestine, hepatic lesions may be uncommon. An atypical enteric syndrome has been described in H. muridarum-infected mice with gastritis and gastric atrophy, but Helicobacter spp. are more typically associated with lower bowel disease in mice.
Hepatic lesions are quite variable. If present, they consist of randomly scattered grossly visible white foci up to 4 mm in diameter. Early lesions may be confined to one or more lobes of the liver, with focal necrosis and mixed leukocytic infiltrates (Fig. 1.60). Within a few months, there is marked hypertrophy and hyperplasia
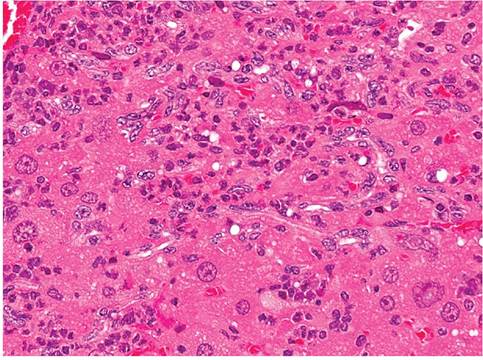
FIG. 1.61. Liver from a mouse chronically infected with Helicobacter hepaticus. Note the hepatocytomegaly, polykarya, Ito cell proliferation, and leukocytic infiltration.
of Kupffer, Ito, and oval cells, with increased mitotic activity among hepatocytes. There may be prominent bile ductule formation extending from the portal regions with apoptosis of individual hepatocytes. Cellular infiltrates consist primarily of lymphocytes and plasma cells (Fig. 1.61). In typical cases of chronic hepatitis, the elongated, helical microorganisms are best demonstrated within biliary canaliculi using the Steiner silver staining method (Fig. 1.62). Selected strains of mice, such as A/J mice, with Helicobacter-associated hepatitis develop foci of cellular alteration, including clear cell, vacuolated, and basophilic foci, with an increased incidence and earlier onset of hepatocellular tumors. C57L mice infected with H. bilis and fed a lithogenic diet develop a high frequency of gallstones.
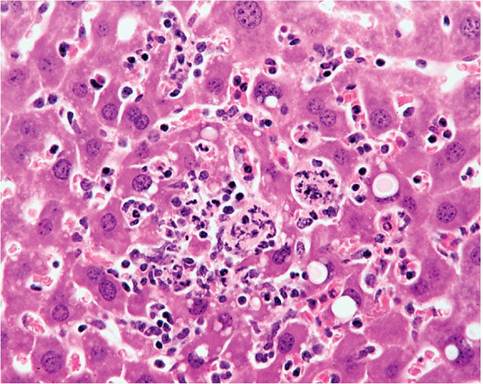
FIG. 1.60. Liver from a mouse infected with Helicobacter hepaticus with focal necrotizing hepatitis.
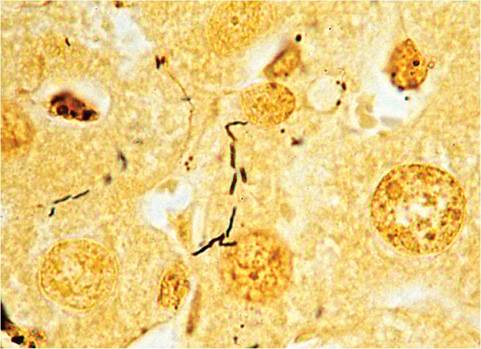
FIG. 1.62. Liver from a mouse infected with Helicobacter hepaticus depicting characteristic Helicobacter organisms within bile canaliculi (Steiner stain).
Diagnosis
Diagnosis is based upon the liver or intestinal lesions, but other etiologic agents must be considered. Definitive diagnosis requires culture, PCR, or demonstration of typical organisms within bile canaliculi or crypts using silver stains. The major challenge to serologic or molecular methods of diagnosis is the antigenic and genetic diversity among the various species of mouse Helicobacter. Major impediments to successful serology are the fact that there are no known or useful cross-species Helicobacter antigens. Furthermore, immunocompetent mice mount minimal antibody responses, and generally only late in infection. Relatively insensitive serologic assays are available that utilize species-specific membrane antigen extracts or recombinant proteins for detecting serum IgG and fecal IgA. PCR amplification of a conserved segment of 16S rRNA gene can be used to detect most Helicobacter spp., and embellished with restriction enzyme analysis to speciate Helicobacter. Multiplex species-specific PCR primers have also been developed for discrimination of several species. PCR is frequently used to test feces, but the authors have found that some species of mouse Helicobacter may not be consistently shed in feces, or only intermittently so. Optimal PCR detection (and culture) can be achieved by sampling cecal mucosa. Differential diagnosis of liver lesions includes infections with Salmonella, Proteus, C. piliforme, MHV, ectromelia virus, or nonspecific foci of hepatic necrosis and inflammation. Intestinal lesions must be differentiated from those induced by infection with E. coli, C. rodentium, enterotropic MHV, or the nonspecific lesions of rectal prolapse.
More on the topic Helicobacter spp. Infections:
- Helicobacter spp. Infections
- Using PCR and 16S rRNA sequence analysis, gastric biopsy specimens were evaluated for the presence of Helicobacter spp. among pet, laboratory, and commercial rabbits. Rabbits from all sources tested positive for Helicobacter spp. Most of th
- Helicobacter spp. Infection
- HELICOBACTER INFECTIONS
- Other Bacterial Infections Corynebacterium spp. Infections
- Mycobacterium spp. Infections
- Staphylococcus spp. Infections
- Bacterial Enteric Infections Brachyspira spp. Infection
- Cryptosporidium spp. Infections
- Mycoplasma spp. Infections
- Malassezia spp. Infection: Malasseziasis
- Aspergillus spp. Infection
- Actinobacillus spp. Infection
- Actinomyces spp. Infection
- Leptospira spp. Infection
- Aspergillus spp. Infection: Aspergillosis
- Brucella spp. Infection: Brucellosis
- Klebsiella spp. Infection