Parvovirus Infections
Laboratory rats are naturally susceptible to 4 serotypes of parvovirus: rat virus (RV, or Kilham's rat virus), H-1 virus (or Toolan's H-1 virus), rat parvovirus (RPV), and rat minute virus (RMV).
RV, H-1, and RMV are antigenically and genetically closely related, whereas RPV is more disparate. Each group contains a number of isolates.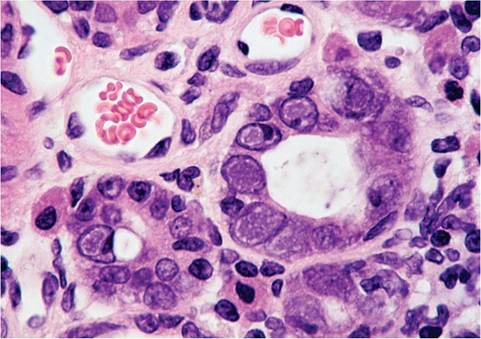
FIG. 2.6. Submandibular salivary gland of a nude rat naturally infected with rat polyomavirus. Note the multiple prominent intranuclear inclusions with margination of nuclear chromatin.
Epizootiology and Pathogenesis
Serological surveys indicate that subclinical infections with various rat parvoviruses are common among laboratory rats. Transmission appears to occur primarily by oronasal contact with infected animals, or by contaminated fomites. Virus shedding has been documented in urine, feces, and oropharynx. RV is recognized to be the most pathogenic of rat parvoviruses under both field and experimental conditions and may be the only strain that produces clinical disease under natural conditions. Transplacental transmission of RV has been demonstrated in pregnant rats inoculated orally with high doses of RV, resulting in infertility and fetal resorption. RV may persist in colonies for long periods of time. Rats born to seropositive dams receive maternal antibodies during the neonatal period and usually acquire RV at 2-7 months of age. Virus may also be shed in the milk during lactation. In seronegative 2-day-old suckling rats inoculated orona- sally with RV, inoculated animals were able to transmit virus for up to 10 weeks, and for at least 7 weeks after seroconversion had occurred. Rats inoculated as juveniles shed RV for at least 3 weeks postinoculation. Virus may be reactivated by immunosuppression, resulting in acute systemic disease.
Although persistence can theoretically occur in individual rats, persistence of parvovirus infections in colonies of rats is dependent on the continuous availability of new susceptible animals to permit propagation of the virus. The widespread distribution of parvoviruses and their requirements for dividing cells for replication results in the frequent contamination of tumor cell lines and tumor virus stocks that have been passaged in laboratory rats.In experimentally induced infections in newborn rats, target tissues for RV replication include primordial cells of the cerebellar cortex, periventricular region, hepatocytes, endothelial cells, and bone marrow. Other target organs include kidney, lung, and genital tract. The multiple hemorrhages seen in the experimental and naturally occurring disease are attributed to the endothelial cell and megakaryocyte damage associated with viral replication in these tissues. Intestinal mucosal lesions, which are so prominent in feline and canine parvoviral infections, do not occur in rats, which may be due to a lack of viral receptors on enterocytes. Documented descriptions of spontaneous outbreaks of parvovirus disease in rats are rare. In 1 report, juvenile rats were clinically affected. Clinical signs included dyspnea, ruffled hair coat, muscular weakness, and cyanotic scrotums. In this outbreak, rats seroconverted to RV, and later to H-1, suggesting that H-1 may have potentiated the development of the disease. Naturally occurring disease has not been attributed to H-1 viruses or RPVs. Experimental studies indicate that both of these types of parvovirus are also prone to persistence, and RPV has a strong tropism for lymphoid tissue, which may cause immunomodulation in experimental studies.
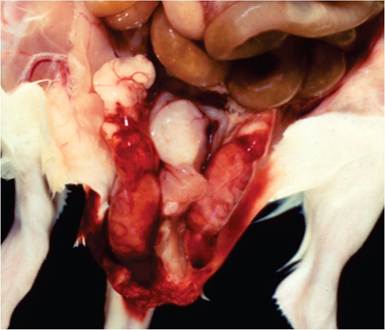
FIG. 2.7. Rat virus (RV) infection in a young rat. Note the acute hemorrhage and fibrinous exudation in the peritesticular region.
(Source: Coleman, et al. 1983. Reproduced with permission from SAGE Publications.)Pathology
In the adult form of RV disease, there is congestion of lymph nodes, loss of body fat, and scrotal hemorrhage, with peri-testicular fibrinous exudation (Fig. 2.7). Splenomegaly, icterus, and ascites are variable findings. Microscopic changes may be present in the brain, liver, and testes. Disseminated foci of hemorrhage occur in the cerebrum and cerebellum in a random distribution involving gray and white matter with malacia and obliteration of the normal architecture (Fig. 2.8). In the testes and epididymis, there may be multifocal coagulation necrosis and hemorrhage consistent with infarction, with thrombosis of regional vessels. Focal hepatocellular necrosis may occur, and amphophilic, intranuclear inclusions may be present in hepatocytes (Fig. 2.9), endothelial cells, and bile duct
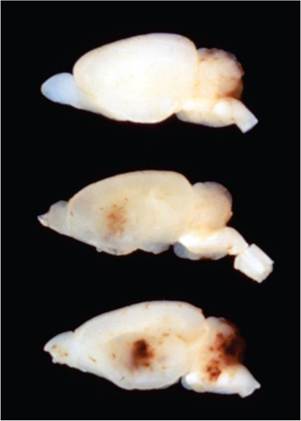
FIG. 2.8. Brains of juvenile rats experimentally infected with RV, depicting hemorrhagic encephalopathy. (Source: R.O. Jacoby. Yale University, New Haven, CT. Reproduced with permission from R.O. Jacoby.)
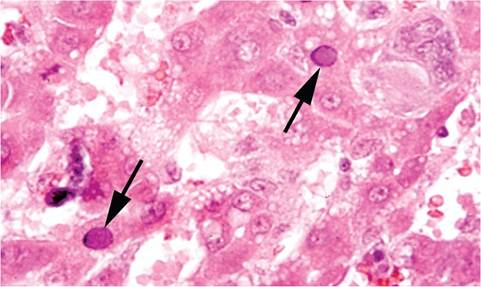
FIG. 2.9. Liver from a juvenile rat with RV infection, illustrating intranuclear inclusions (arrows) in hepatocytes. (Source: R.O. Jacoby. Yale University, New Haven, CT. Reproduced with permission from R.O. Jacoby.)
epithelium. Neonatal or infant rats can develop granulo- prival cerebellar hypoplasia, hepatitis, and jaundice. Lesions present in recovered rats may include focal angiec- tasis (peliosis hepatis), and nodular hyperplasia with portal scarring. Infertility, fetal resorption, and abortion may occur in pregnant females.
Diagnosis
The provisional diagnosis, based on the presence of typical lesions and inclusions, can be confirmed by the demonstration of viral antigen by immunohistochemistry or detection of DNA by PCR. Seroconversion may be detected by various procedures, but contemporary methods now utilize recombinant VP2 and NS1 antigens. VP antigens reflect virus strain specificity, whereas NS antigens are highly conserved and cross-reactive among all of the rat (and mouse) parvoviruses. RPV does not share homologous cross-reacting structural VP antigens with RV, H-1, or MVR, but possesses conserved cross-reacting NS nonstructural antigens. Likewise, PCR assays may target either NS1 or VP2 gene sequences. PCR has been used to detect rodent parvoviruses in contaminated biomaterials, mesenteric lymph node, spleen, feces, and environmental surfaces. Differential diagnoses include bacterial septicemias, chronic wasting due to agents such as Mycoplasma pulmonis, and trauma. Infertility and fetal resorption must be differentiated from conditions such as nutritional disorders, disruption of normal light cycles, and mycoplasmal infections.
More on the topic Parvovirus Infections:
- OTHER PARVOVIRUS INFECTIONS
- CHAPTER 12 PARVOVIRUS INFECTIONS
- PORCINE PARVOVIRUS INFECTION IN WILD BOAR
- Parvovirus Infection
- Other Bacterial Infections Corynebacterium spp. Infections
- Bloodstream Infections and Catheter-Related Bloodstream Infections
- Skin, Soft-Tissue, and Bone Infections Purulent Skin and Soft-Tissue Infections (Furuncles, Carbuncles, Abscesses)
- OPPORTUNISTIC INFECTIONS
- INTRAUTERINE INFECTIONS
- PERINATAL INFECTIONS
- NOSOCOMIAL INFECTIONS
- INTRACRANIAL INFECTIONS
- ENTERQViRAL INFECTIONS (NON-POLIO)
- Genitourinary Infections
- HERPETIC VIRAL INFECTIONS
- FUNGAL INFECTIONS
- Viral infections
- Tick-Borne Infections
- 10.10 MENINGOCOCCAL INFECTIONS
- Bioterrorism and Emerging Infections