POLYMORPHUS/PROFILICOLLIS SPP.
CAUSATIVE AGENT (CLASSIFICATION, morphology) There is some ambiguity on
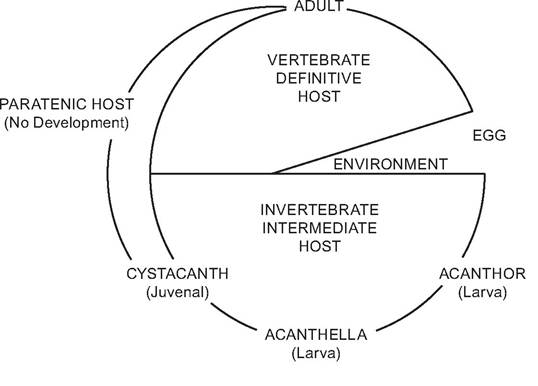
FIGURE 3.11 Diagram of the basic acanthocephalan life cycle (DeGiusti 1971; with permission of John Wiley & Sons, Inc.).
the status of the genus, Polymorphus (Order Polymorphida, Class Palaeacanthocephala). At one time it was a broad genus with at least 35 species and two subgenera, Polymorphus and Profilicollis (Amin 1985). More recently the two subgenera have been proposed as independent genera (Nickol et al. 1999); however, much of the wildlife literature is based on the older and broader taxonomy. The larger Polymorphus group will be addressed in this section, but the two genera (former subgenera) will be specifically named when this is evident. Members of this group commonly have been associated with mortality among waterfowl and other birds. One of the more commonly observed species is Polymorphus minutus, which affects a number of families of birds, including sea ducks and other waterfowl (Janovy 1997). Profilicollis botulus often has been associated with mortality in common eiders (Somateria mollissima) (Thieltges et al. 2006).
host range and distribution Members of these two genera have been reported from all parts of the world. Most reports come from aquatic birds, although a few species occur in mammals, including marine mammals (Crompton and Nickol 1985)
life cycles and variations All members of Profilicollis spp. use crabs of the crustacean order Decapoda as intermediate hosts (Nickol et al. 1999). For Profilicollis botulus in northern Europe, the littoral crab (Carcinus maenas) (Portunidae) is the principal intermediate host, with the crab distribution and habitat overlapping with its definitive host, eider ducks (Somateria mollissima) (Nickol et al.
1999); P. botulus also is found in American lobsters (Homarus americanus) (Nephrodpidae) of the maritime provinces of Canada (Brattey et al. 1985).Members of the revised Polymorphus spp. commonly use amphipods. Polymorphus minu- tus uses ducks as definitive hosts and freshwater amphipods such as Gammarus pulex as intermediate hosts (Crofton 1971).
In both genera, acanthocephala eggs are passed from the adults in the definitive host into the aquatic environment. After ingestion by an appropriate intermediate host, they develop the acanthor, acanthella, and cystacanth stages in their respective intermediate hosts, which, in turn, are ingested by the definitive host. The cystacanths excyst and adult worms attach to the small intestine.
reservoirs and transmission Polymor- phus minutus can survive 15 months as a cystacanth in its amphipod intermediate host and 1.7 months in the duck definitive host (Crompton and Whitfield 1968). Profilicollis botulus can survive 36 months as a cystacanth in the decapod intermediate host and 3.3 months as an adult in the definitive host (Thompson 1985a, 1985c).
Transmission to the definitive host can be enhanced by making the infected intermediate host more visible or more attractive to the definitive host. Amphipods (Gammarus Iacustris) infected with Polymorphus spp., probably P. minutus, were a blue color in contrast to the normal gray-to-brown color of uninfected amphipods (Hindsbo 1972). Based on laboratory experiments on other species (e.g., Polymorphus paradoxus, Polymorphus marilis), amphipods containing cystacanths infective for ducks preferred being in lighted areas or were especially sensitive to disturbance and spent more time at the surface of the water; this was in contrast to uninfected amphipods. The changed behaviors resulted in ducks selecting many more acanthocephala-infected amphipods (Bethel and Holmes 1973, 1977; Moore 2002; Kennedy 2006).
CLINICAL EFFECTS AND IDENTIFICATION In laboratory studies, Polymorphus-infected common eider ducklings gained weight more slowly than control birds, and had lower serum concentrations of total protein, β-globulin, g-globulin, fructosaminase, and creatine kinase; parasites were attached to the mucosa of the posterior small intestine, with a mixed inflammatory reaction consisting of hetero - phils and lymphocytes around the attachment sites (Hollmen et al.
1999). Polymorphus spp. also causes peritonitis and emaciation in sea ducks (Skerratt et al. 2005). These parasites also can cause cachexia, with heavy losses occasionally causing mortality (Soulsby 1968).There are not many well-documented cases of acanthocephala causing significant mortality in wildlife. More likely, these parasites probably add an additional energy and physiologic burden for hosts and may also interfere with feeding success (Thieltges et al. 2006), with greater impact during periods of food shortage or other environmental stresses.
Fecal evaluations can be used to verify the presence of infection, but do not provide a definitive diagnosis of disease; postmortem examinations are required to reach a diagnosis on the cause of death (Cole 1999). Attributing mortality to these acanthocephala should be based on associated pathology rather than the numbers of parasites, since sea ducks may harbor large numbers with no obvious clinical effects (Garden et al. 1964, Thompson 1985b, Camphuysen et al. 2002).
population effects There are a number of reports of species of Polymorphus/Profilicollis causing mortality in wild sea ducks and other birds (MacDonald et al. 1978, Skerratt et al. 2005). Heavy burdens of Profilicollis altmani caused peritonitis and possible emaciation and were associated with an estimated mortality of about 100 surf scoters (Melanitta perspicil- lata) near San Francisco, California, in 1995 (Skerratt et al. 2005). There have been reports of mortality of Profilicollis botulus among common eiders (Somateria mollissima), which is addressed next.
special problems Common eiders have experienced repeated and sometimes severe mortalities in northern and western Europe; many of these events are associated with Pro- filicollis botulus (Clark et al. 1958, Swennen and VanDenBroek 1960, Rayski and Garden 1961, Grenquist 1970, Persson 1974, Morner 1978, Itamies et al. 1980), and also, on occasion, with Polymorphus minutus (Christiansen 1948, Itamies et al.
1980). Among common eiders, mortality often occurs in the late winter among immature birds and adult males, whereas adult female mortality is associated with the time of brooding (Garden et al. 1964, Grenquist 1970).Common eiders generally have a rich endoparasite fauna, in particular with the acanthoceph- alan Profilicollis botulus (Thieltges et al. 2006). Prevalence typically is similar among both young and adult birds, but the young have far greater intensities of P. botulus (Garden et al. 1964, Liat and Pike 1980, Camphuysen et al. 2002). It is not clear whether these differences are due to differences in susceptibility by age, an acquired immunity that aids adults, differential food source use leading to different exposures, or high mortality among juveniles with high intensities and resulting lower intensities among surviving adults (Thieltges et al. 2006).
There is some uncertainty on the role of Profilicollis botulus on eiders. Healthy eiders virtually always have some infections with acan- thocephala, often with large numbers (Garden et al. 1964, Bishop and Threlfall 1974, Persson et al. 1974, Liat and Pike 1980, Thompson 1985b, Camphuysen et al. 2002). In one review of acan- thocephalan outbreaks, there was no evidence for direct parasite-induced mortality in common eiders, but rather evidence for a combined occurrence of a weakened physical condition and heavy parasite intensity (Hario et al. 1995).
In a mass mortality event involving about 21,000 common eider ducks observed in the southwestern part of the Wadden Sea in the winter of 1999/2000 in the Netherlands and Germany (Camphuysen et al. 2002), acantho- cephala and food availability were considered to be significant contributors to this event (Borgsteede 2001). While there was some controversy on the relative contributions role of food availability and parasites in that mortality event (Smaal et al. 2001, Camphuysen et al. 2002), the more common perspective was that the parasite infections, while extensive, likely were not directly responsible for the observed mortality (Camphuysen et al.
2002, Thieltges et al. 2006).There is a possibility that this mortality followed a combination of events and circumstances. Winter strength, food depletion, and other stressors in combination with high parasite loads can result in mortality (Hario et al. 1995). The high numbers of parasites at the Wadden Sea event may have depleted the energy reserves of the eiders or interfered with their successful feeding. Because P. botulus can induce severe lesions in the intestines, the eiders might have switched from hard-shelled molluscs to softer prey such as crabs, in turn leading to increased acanthocephalan infections (Thieltges et al. 2006). With an increasing parasite load, feeding ability could have been further compromised, leading to malnutrition and even starvation (Thieltges et al. 2006). At this point, the role of acanthocephala still is not clear on this issue.
control No control has been proposed for acanthocephala among free-living wildlife. Carbon tetrachloride at a dose of 0.5 ml per kg has been reported to be up to 98% effective among domestic animals (Soulsby 1968). Thiabendazole has been recommended for birds, but treatment is acknowledged to be difficult and is generally not successful (Cole 1999).
Other Acanthocephala
There are a number of other acanthocephala of importance to wildlife. Among species infecting birds, one example is Plagiorhynchus cylindraceus, which uses European starlings (Sturnus vulgaris) and other songbirds as definitive hosts, and terrestrial isopods (e.g., Armadil- lidium vulgare, “pillbug”) as intermediate hosts (Schmidt 1985, Moore 2002). This is a case in which the acanthocephalan changes the behavior of the infected intermediate host to make it spend more time in areas of reduced humidity, and to spend more time on light than dark surfaces; such behavioral changes probably make the infected pillbugs more conspicuous to visual predators and are associated with a disproportionate predation on infected pillbugs (Moore 1983, 1984).
However, the impacts of adult P. cylindraceus on the starling usually are minor (Moore and Bell 1983, Connors and Nickol 1991).Among mammalian acanthocephala, members of the genera Macracanthorhynchus and Moniliformis are considered potentially significant in wildlife (DeGiusti 1971). Macra- canthorhynchus hirudinaceus is the giant thorny- headed worm with a worldwide distribution, commonly found in domestic pigs; the parasite also has been reported from many species of wildlife, including feral and wild suids, ground squirrels, tree squirrels, muskrats, moles, hyenas, and other species (DeGiusti 1971). Adults occur in the small intestine of the definitive host; intermediate hosts include many species of beetles of the Family Scarabaeidae and some other Coleoptera (DeGiusti 1971, Schmidt 1985). No paratenic hosts are reported (Schmidt 1985). While mortality is not common, pathology can include an acute necrotizing enteritis and perforation of the intestinal wall with ensuing peritonitis and occasional death of the host (DeGiusti 1971).
Moniliformis moniliformis also has a cosmopolitan distribution, and is found in a wide variety of rodents, as well as lagomorphs and several other mammalian species (DeGiusti 1971). Intermediate hosts include roaches and a variety of beetles (DeGiusti 1971, Schmidt 1985). Parasite-infected intermediate hosts in some species appear to lose interest in breeding and are “behaviorally castrated” (Carmichael et al. 1993). Interestingly, some arthropod intermediate hosts appear to retard development of the parasite within them by selecting cooler temperatures (Moore and Freehling 2002).
More on the topic POLYMORPHUS/PROFILICOLLIS SPP.:
- Using PCR and 16S rRNA sequence analysis, gastric biopsy specimens were evaluated for the presence of Helicobacter spp. among pet, laboratory, and commercial rabbits. Rabbits from all sources tested positive for Helicobacter spp. Most of th
- Malassezia spp. Infection: Malasseziasis
- Helicobacter spp. Infections
- Bacterial Enteric Infections Brachyspira spp. Infection
- Actinobacillus spp. Infection
- Actinomyces spp. Infection
- Staphylococcus spp. Infections
- Helicobacter spp. Infections
- Leptospira spp. Infection
- Helicobacter spp. Infection
- Aspergillus spp. Infection: Aspergillosis
- Brucella spp. Infection: Brucellosis
- Enterococcus spp. Infection: Enterococcal Enteropathy
- OTHER ECHINOCOCCUS SPP.
- Dirofilaria spp. Infestation
- Chlamydophlla spp. Infection
- Klebsiella spp. Infection
- Cryptosporidium spp. Infections